



第二节 生物氧化
所有生物体在生命活动过程中都需要能量,能量的来源依靠生物体不断从外界摄取营养物质或有机物质(糖、脂肪、蛋白质等)在体内氧化来供给,食物中的糖、脂肪、蛋白质等有机物,通常称为人体的三大营养素,那么什么是生物氧化呢?
一、生物氧化的概念、意义及特点
1.生物氧化的概念和意义
生物氧化就是指生物从外界摄取的有机物质,在生物细胞内氧化分解,最终彻底氧化分解成二氧化碳和水,并释放出能量的过程。由于生物氧化是在组织细胞中进行的,氧化过程又与吸人氧和呼出二氧化碳的呼吸作用密切相关,故又将生物氧化称为细胞氧化或细胞呼吸。
氧化还原的本质是电子的转移,在生物体内,氧化的形式有以下几种:
(1)直接进行电子转移
Fe2++Cu←↑2+Fe3++Cu+
(2)氢原子的转移 因为H原子可分解成为H+和e-,因此其本质也是电子转移。
AH2←↑+BA+BH2
(3)有机物加氧 因加氧时常伴有氧接受质子和电子而被还原成水,其本质也是电子转移。
RH+O2+2H++2e←↑-ROH+H2O
生物氧化的意义就在于它能为机体提供生命活动所需的能量。
2.生物氧化的特点
生物氧化与体外物质氧化或燃烧的化学本质是相同的,即都是消耗氧,使有机物氧化,最终生成二氧化碳和水,释放出的总能量也相等,但是有机物的生物氧化又有其特点。
生物氧化是在细胞内进行,是在体温和近于中性pH值及有水环境中进行的,是在一系列酶、辅酶和中间传递体的作用下逐步进行的,每一步都放出一部分能量,这样不会因温度迅速上升而损害机体,使释放的能量得到有效的利用。生物氧化释放的能量通常都先贮存在一些特殊的高能化合物(如ATP等)中,在需要的时候再由ATP分子释出,并可以转换成各种形式的能量,以供机体生命活动的需要。
真核细胞的生物氧化是在线粒体中进行,而原核生物细胞的氧化是在细胞质膜上进行。
有机物在生物细胞内是如何被彻底氧化分解成二氧化碳和水,并释放出能量的过程将在糖、脂肪和氨基酸代谢中介绍,生物氧化主要讨论的问题:一是细胞如何利用O2将代谢物分子中的氢氧化成水;二是代谢物中碳如何在酶催化下生成CO2;三是有机物被氧化时产生的自由能如何被收集、贮存或利用。
二、生物氧化中二氧化碳的生成
生物氧化中生成的CO2是由于糖、脂类、蛋白质等有机物转变成含羧基的有机酸进行脱羧反应所致。根据所脱羧基在有机酸分子中所处位置分为α-脱羧和β-脱羧。脱羧过程伴随氧化作用的称为氧化脱羧,没有氧化作用的称为直接脱羧。α-直接脱羧,如丙酮酸脱羧,其反应式如下:
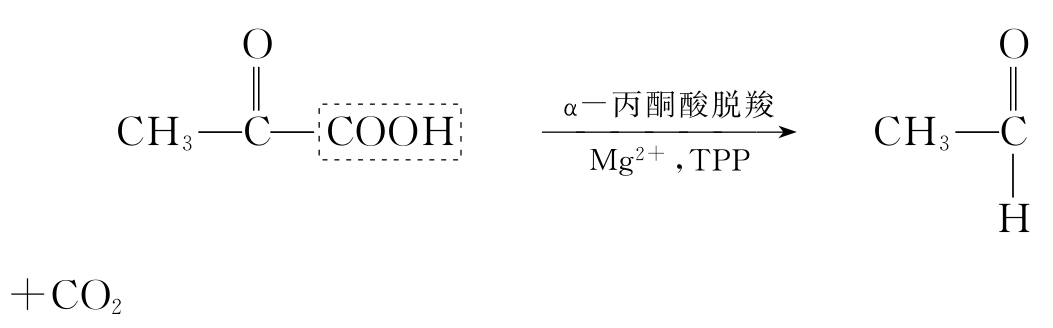
β-直接脱羧,如草酰乙酸脱羧,反应式如下:
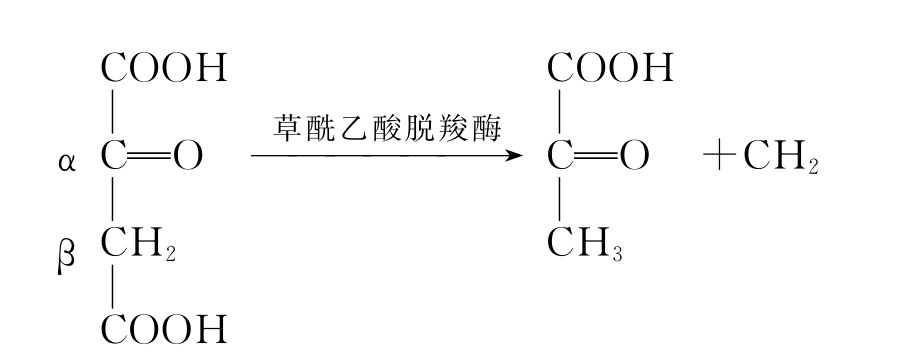
α-氧化脱羧,如丙酮酸的氧化脱羧,见如下反应:
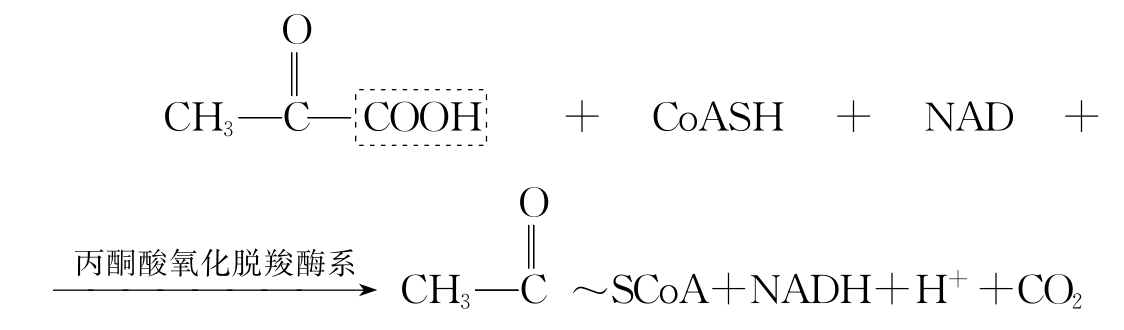
β-氧化脱羧,如苹果酸的氧化脱羧,见如下反应:
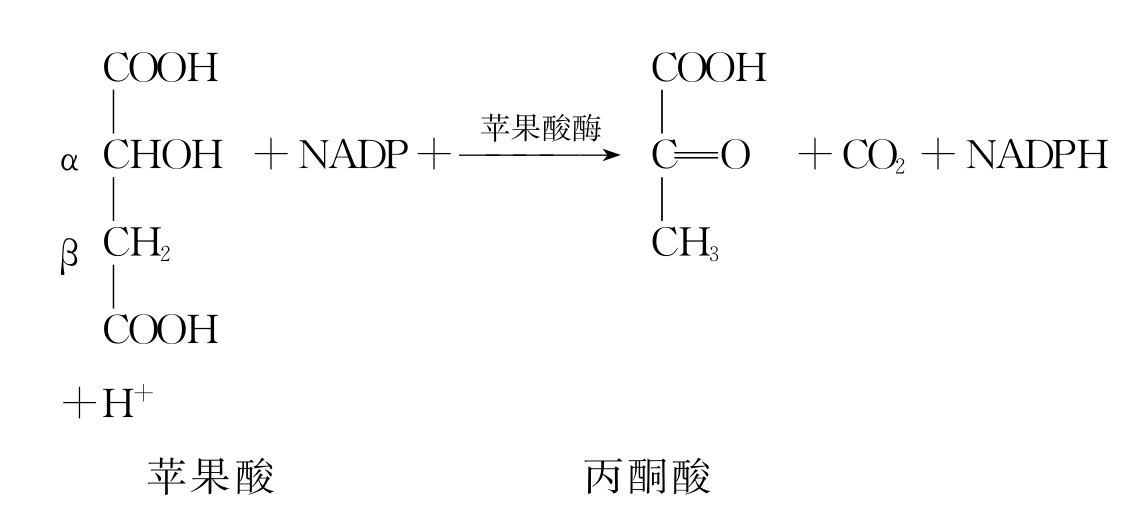
三、生物氧化中水的生成
H2O是生物氧化的产物之一,脱氢是氧化的一种方式,生物氧化中生成的水是代谢物脱下的氢,经过一系列传递体的传递,最后与吸人的氧结合生成水(图7-1),所以生物氧化是需氧的过程。生物体中有机物所含的氢,一般情况下是不活泼的,必须通过相应的脱氢酶将之激活后才能脱落;进人体内的氧也必须经过氧化酶激活后才能变为活性很高的氧化剂。脱落的氢需经过传递体传递才能与氧结合生成水。所以,生物体内的生物氧化体系主要是由脱氢酶、氧化酶和传递体组成的。
四、生物氧化中能量的产生(线粒体氧化体系)
生物氧化的最终结果是生成二氧化碳和水,同时伴随着自由能的释放和ATP的生成。对于真核生物,真核细胞的线粒体是能量转换的重要部位,参与生物氧化的酶、传递体主要存在于线粒体中,所以也把生物氧化能量的产生称为线粒体氧化体系。在能量的产生中主要涉及参与氧化的酶类、呼吸链和能量的产生机制——氧化磷酸化作用。
1.线粒体
线粒体是一种较大的细胞器。线粒体具有双层膜结构,外膜平滑,透性高,大多数小分子化合物(相对分子质量在10000以下的物质)均能通过,仅有少量酶结合在其上。内膜则不同,具有很多向内折叠的嵴[图7—2(a)]。在内膜上包含参与电子传递和氧化磷酸化的有关组分,是线粒体功能的主要担负者,产能和需能越高的组织如翅肌,嵴的数目也越多。线粒体的内膜表面上分布有许多排列规则的球状颗粒,通过短柄与内膜相连[图7-2(b)],这就是ATP合成酶复合物Fo、FI因子。线粒体的内腔充满半流动的基质,其中包含大量的酶类,如与三羧酸循环、脂肪酸β-氧化和氨基酸分解代谢有关的酶都存在于基质中。除此以外,基质中还含有DNA和核糖体,大多数哺乳动物的线粒体DNA为环状分子,线粒体DNA可编码Fo的疏水亚基、细胞色素氧化酶和细胞色素b复合物等,约占内膜总蛋白质的20%左右,其余的蛋白质均由核基因编码,在细胞质中合成后送入线粒体中。
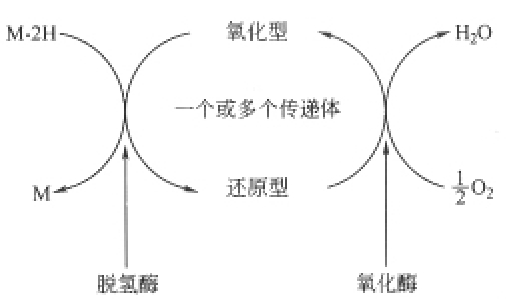
图7-1 生物氧化体系示意
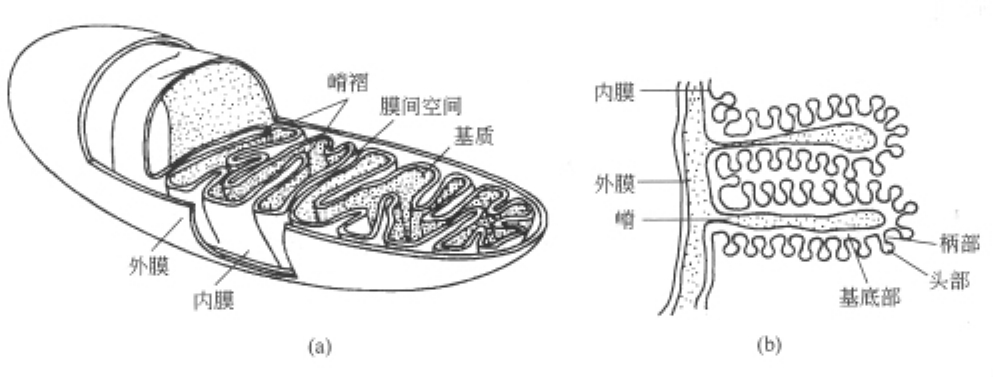
图7-2 线粒体结构示意
2.参与生物氧化的酶类
(1)氧化酶类——氧的还原 氧化酶是一类含金属离子(如钼、铁、铜等)的结合酶。其共同特点是:直接以氧作为受氢体,所催化的反应无氧不能进行;分子氧在氧化酶的催化下,每个氧原子接受2个电子(2e-)后和2个质子(2H+)化合成H2O;酶分子中的金属离子作为电子的传递体。其作用方式如下式所示:
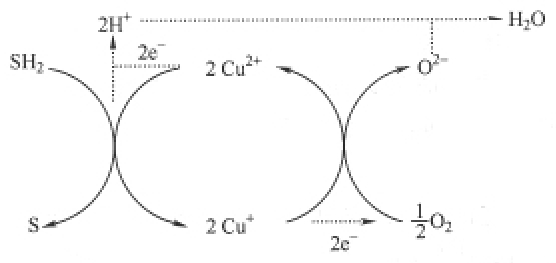
式中,SH2为底物;S为产物。
这类酶易被氰化物、硫化氢、一氧化碳等物质所抑制而失去催化作用。常见的氧化酶类有细胞色素氧化酶类、抗坏血酸氧化酶类和酚氧化酶类等。在线粒体氧化体系中重要的是细胞色素氧化酶类。
(2)脱氢酶类 能使代谢物的氢活化、脱落并将其传递给受氢体或中间传递体的一类酶称为脱氢酶。根据所含辅因子的不同,可将脱氢酶分为两类。
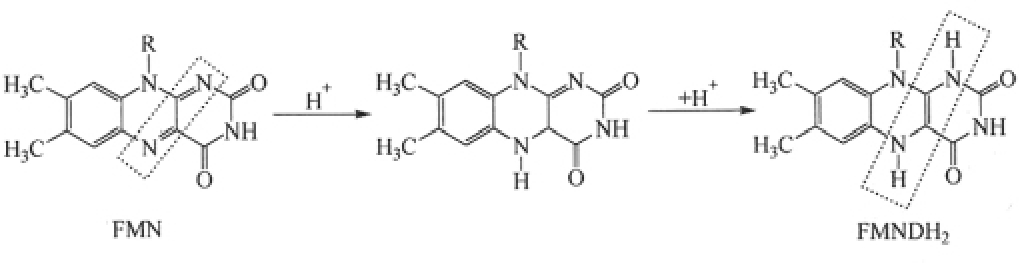
第一类,以黄素核苷酸为辅基的脱氢酶。这类酶以黄素单核苷酸(FMN)或黄素腺嘌呤二核苷酸(FAD)为辅基,又称黄酶。根据最终受氢体的不同,还可将黄酶分为以下两类:
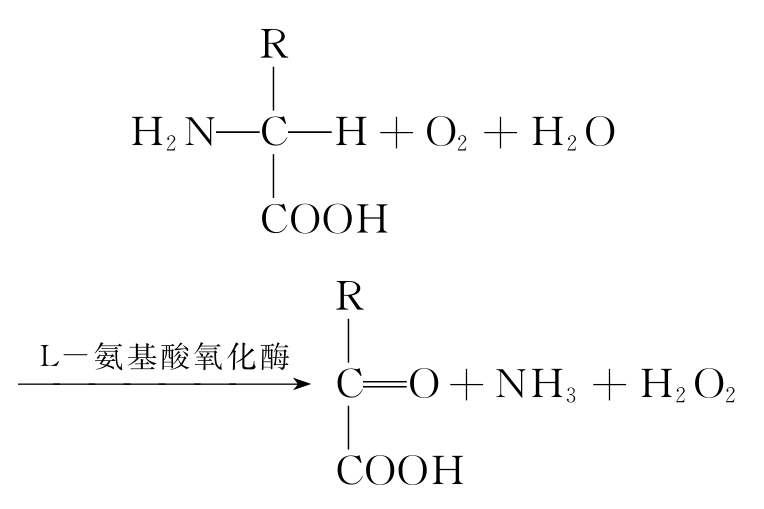
①需氧黄酶 以氧为直接受氢体的称为需氧黄酶,其氧化底物脱下的氢由氧接受产生过氧化氢,如L-氨基酸氧化酶反应过程如下:
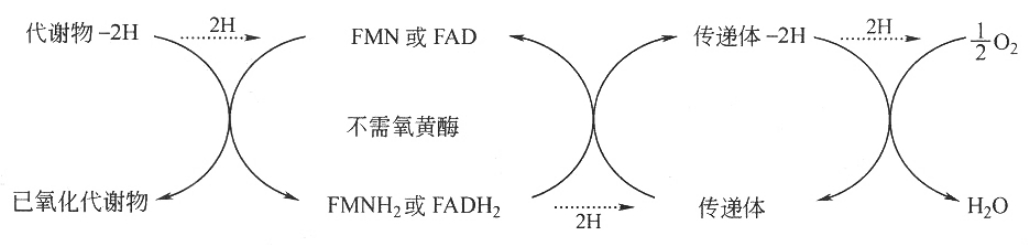
②不需氧黄酶 不以氧为直接受氢体,催化底物脱下的氢先经中间传递体,再传给氧生成水,这一类酶称为不需氧黄酶,如琥珀酸脱氢酶、NADH脱氢酶、脂肪酰CoA脱氢酶、α-磷酸甘油脱氢酶等都是不需氧黄酶,催化如下反应:
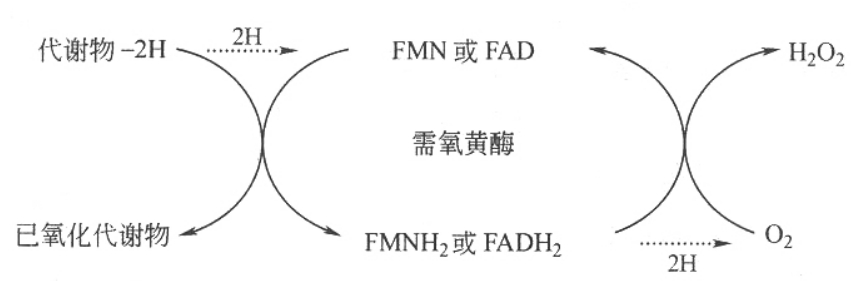
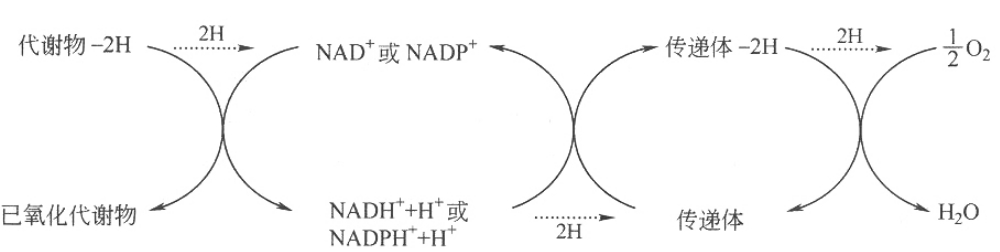
第二类,以烟酰胺核苷酸为辅酶的脱氢酶。此类脱氢酶以NAD(CoⅠ)或NADP(CoⅡ)为辅酶,催化代谢物脱下的氢由NAD+或NADP+接受,再交给中间传递体,最后传给氧生成H2O。这类酶不能以氧为直接受氢体,属于不需氧脱氢酶。催化反应如下:
(3)传递体 在生物氧化过程中起传递氢或传递电子作用的物质称为传递体,传递体既不能使代谢物脱氢,也不能使氧活化。能传递氢原子的传递体称为递氢体,如辅酶Q等;能传递电子的传递体称为递电子体,如细胞色素及铁硫蛋白。
3.呼吸链
(1)呼吸链的概念 氧化体系中的酶类、传递体按一定的规律分布、排列在线粒体内膜和基质中,将代谢物脱下的氢传递给氧生成水,由于该过程利用氧,常将这一复杂体系叫做呼吸链或电子传递链。
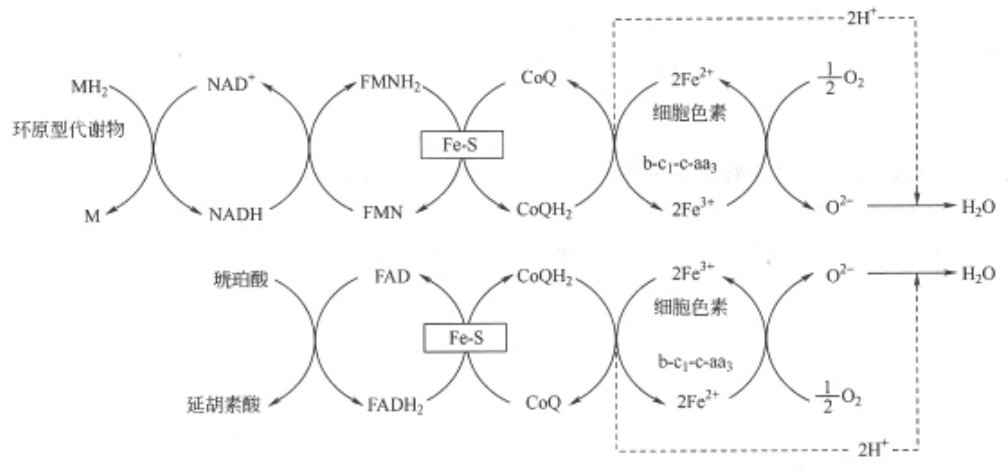
图7-3 NADH、FADH2呼吸链
在具有线粒体的生物中,典型的呼吸链有两种,即NADH呼吸链与FADH2呼吸链(图7—3),这是根据接受代谢物上脱下的氢的初始受体不同区分的。NADH电子传递链分布最广,糖、脂肪、蛋白质这三大物质分解代谢中的脱氢氧化反应,绝大部分是通过NADH呼吸链完成的;FADH2电子传递链中的脱氢酶只能催化某些代谢物脱氢,不能催化NADH或NADPH脱氢,也就是只能作为某些底物的初始受氢体,如琥珀酸的初始受氢体。
在生物体内电子传递链有多种形式,有的是中间传递体的组成不同,例如某些细菌(如分枝杆菌)中的维生素K代替了通常电子传递链中的辅酶Q。形式差异虽大,但电子传递链传递电子的顺序基本上是一致的。生物进化愈高级,呼吸链也就愈完善。
(2)呼吸链的组成 呼吸链是一系列电子传递体按对电子亲和力逐渐升高的顺序组成的电子传递系统,所有组成成分都嵌于线粒体内膜。
目前发现,构成呼吸链的成分有20多种,一般可分为五类,即烟酰胺脱氢酶类、黄素脱氢酶类、铁硫蛋白类、细胞色素类以及辅酶Q类。
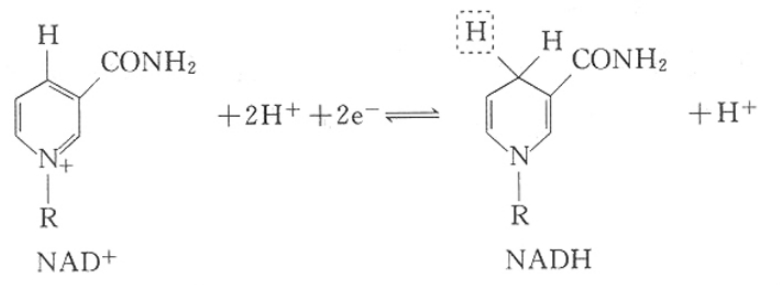
①烟酰胺脱氢酶类 此类酶以含有烟酰胺结构的NAD+或NADP+为辅酶,目前已知者达200多种。此类酶催化脱氢时,其辅酶NAD+或NADP+先和酶的活性部位结合,然后再脱下来。它与代谢物脱下的氢结合而还原成NADH或NADPH。当有受H体存在时,NADH或NADPH上的H可被脱下而氧化为NAD+或NADP+。
NAD++2H(2H++2e-←↑)NADH+H+
NAP++2H(2H++2e-←↑)NADH+H+
②黄素脱氢酶类 以黄素单核苷酸(FMN)或黄素腺嘌呤二核苷酸(FAD)作为辅基。黄素核苷酸与酶蛋白结合是较牢固的。这类酶催化脱氢时现已证明是将代谢物上的一对氢原子直接传给FMN或FAD的异咯嗪基而形成FMNH2或FADH2,异咯嗪基的还原分两步进行。
MH2+酶←↑-FMNM+酶-FMNH2
MH2+酶←↑-FADM+酶-FADH2
③铁硫蛋白类(Fe-S) 铁硫蛋白类的分子中含非卟啉铁与对酸不稳定的硫。其作用是借铁的化合价互变进行电子传递
Fe3++e←↑-Fe2+
因其活性部分含有两个活泼的硫和两个铁原子,故称铁硫中心。
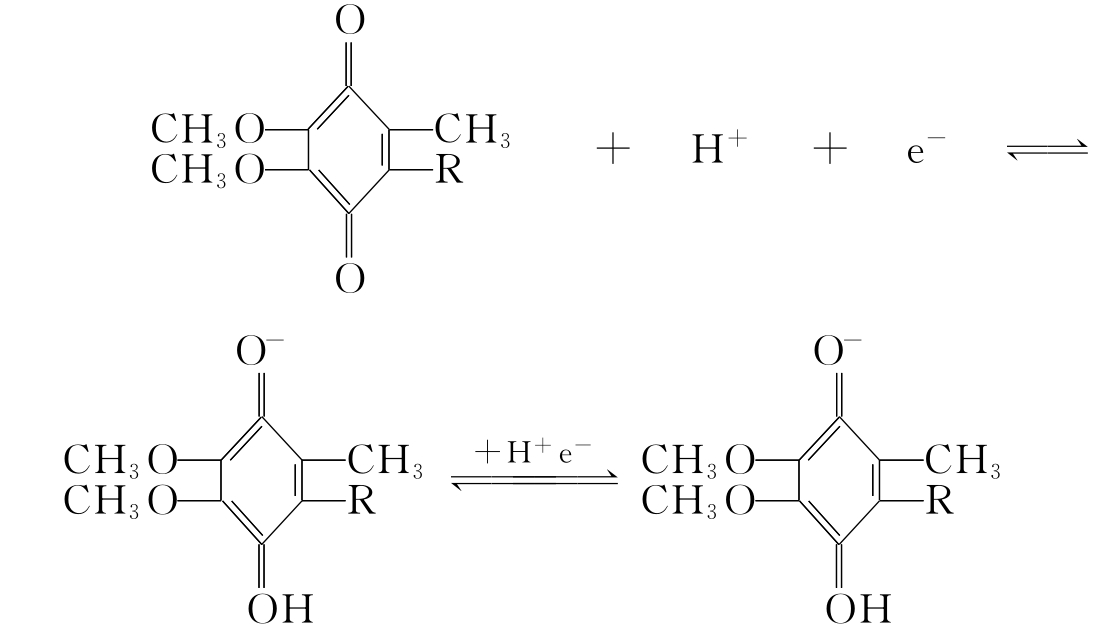
④辅酶Q类 此类酶是一种脂溶性的醌类化合物,因广泛存在于生物界,故又名泛醌。其分子中的苯醌结构能可逆地加氢还原而形成对苯二酚衍生物,故属于传氢体。泛醌可将2个质子释放人线粒体基质内,将电子传递给细胞色素。
⑤细胞色素类 细胞色素是一类以铁卟啉为辅基的蛋白质,在呼吸链中,也依靠铁的化合价的变化而传递电子。
Fe3++e←↑-Fe2+
目前发现的细胞色素有多种,包括a、a3、b、c、c1等,在典型的线粒体呼吸链中,其顺序是b-c1-aa3-O2,其中仅最后一个a3可被分子氧直接氧化,但现在还不能把a和a3分开,故把a和a3合称为细胞色素c氧化酶。
(3)呼吸链中传递体的顺序 呼吸链中的氢和电子的传递是有着严格的顺序和方向的,电子从电子亲和力低(氧化能力弱)的电子传递体传向电子亲和力高(氧化能力强)的传递体,或者说是从低氧化还原电位向高的电位上流动。氧化还原电位E-'的数值愈低,即供电子的倾向愈大,愈易成为还原剂,处于电子传递链的前端。测定各电子传递体的标准氧化还原电位值E-'即可测出其氧化还原能力。
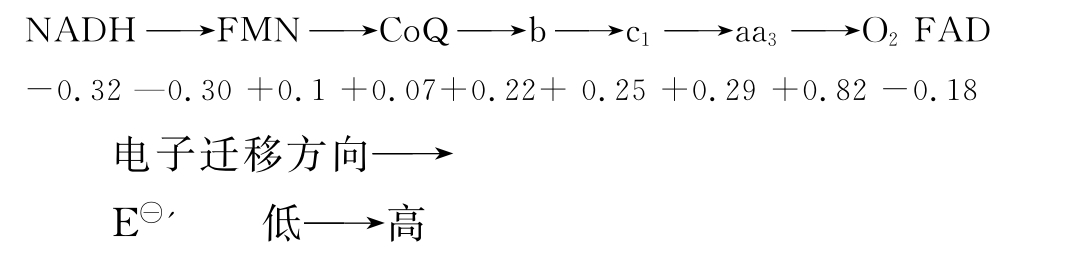
根据 值,细胞色素b应在CoQ之前,但其他实验证明细胞色素b是位于CoQ之后。还有铁硫蛋白的位置问题。这些表明目前呼吸链各成员排列的确顺序尚未弄清,尚有一些细节需进一步确定。
值,细胞色素b应在CoQ之前,但其他实验证明细胞色素b是位于CoQ之后。还有铁硫蛋白的位置问题。这些表明目前呼吸链各成员排列的确顺序尚未弄清,尚有一些细节需进一步确定。
4.氧化磷酸化作用
生物体通过生物氧化所产生的能量,除一部分用于维持体温外,大部分通过磷酸化作用转移至高能磷酸化合物ATP中,这种伴随放能的氧化作用而使ADP磷酸化生成ATP的过程称为氧化磷酸化作用。根据生物氧化的方式,可将氧化磷酸化分为底物水平磷酸化和电子传递体系磷酸化。
(1)ATP的生成 ATP主要由ADP磷酸化生成,少数情况下可由AMP焦磷酸化生成。
ADP+Pi+→─能量ATP
AMP+PPi+→─能量ATP
①底物水平磷酸化 底物水平磷酸化是在被氧化的底物上发生磷酸化作用。即底物被氧化的过程中,形成了某些高能磷酸化合物的中间产物,通过酶的作用可使ADP生成ATP。
x~○→─P+ADP ATP+X
式中,X~○P代表底物在氧化过程中所形成的高能磷酸化合物。
在糖的分解代谢中(如3磷酸甘油醛转变成1,3-二磷酸甘油酸,形成了高能磷酸化合物;α-酮戊二酸氧化脱羧生成琥珀酸),也有高能化合物形成。底物磷酸化形成高能化合物,其能量来源于伴随底物脱氢分子内部能量的重新分布。
底物磷酸化也是捕获能量一种方式,底物磷酸化和氧的存在与否无关,在发酵作用中是进行生物氧化取得能量的惟一方式。
②电子传递体系磷酸化 当电子从NADH或FADH2经过电子传递体系(呼吸链)传递给氧形成水时,同时伴有ADP磷酸化为ATP,这一全过程称电子传递体系磷酸化。电子传递体系磷酸化是生成ATP的一种主要方式,是生物体内能量转移的主要环节。
氧化磷酸化的偶联部位:根据实验证明,从NADH到分子氧的呼吸链中,三处能使氧化还原过程释放的能量转化为ATP,而且这三个释放能量的部位都已弄清。这三处也即是传递链上可被特异性抑制剂切断的地方。NADH呼吸链生成ATP的三个部位是:
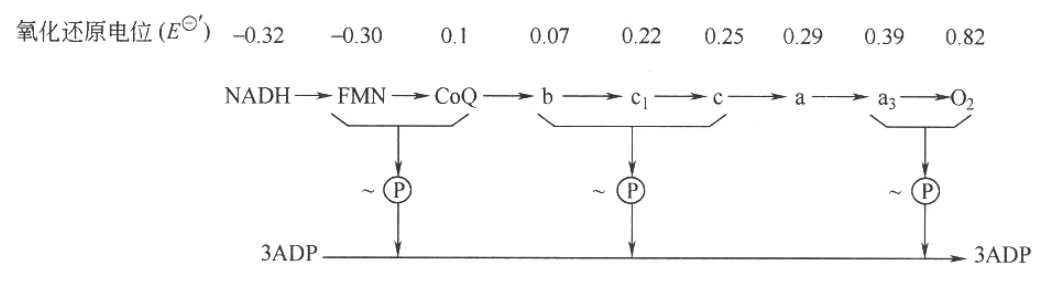
氧化磷酸化的效率可通过测定线粒体的P/O比值来判断。P/O比值是指某一代谢物作呼吸底物,每消耗1mol氧所消耗无机磷酸的物质的量。实验指明NADH呼吸链的P/O值是3,即每消耗1mol氧原子就要消耗3mol的无机磷酸,形成3mol ATP;FADH2链的P/O值是2,即消耗1mol氧原子可形成2mol ATP。从不同底物的氢所得到的P/O值不完全相同,故按不同底物参加呼吸链的P/O比值,可以推断氧化磷酸化的部位。
③影响氧化磷酸化的因素 影响呼吸链的任何因素都影响氧化磷酸化的正常进行。可将这些因素分为三种类型(图7-4):

图7—4 链抑制剂、解偶联剂和离子载体抑制剂的作用部位
a.呼吸链抑制剂 有些物质以专一的结合部位抑制呼吸链的正常传递,影响氧化磷酸化作用,从而妨碍或破坏能量的供给。如阿米妥(麻醉药)、鱼藤酮(杀虫剂)、大黄酸和大黄素等物质抑制NADH—CoQ之间氢的传递;抗霉素A抑制Cy→─tobCytoc之间的电子传递;氰化物、叠氮化物、一氧化碳和硫化氢则抑制细胞色素氧化酶与分子氧之间的电子传递。因此,此类抑制剂可使细胞内呼吸停止,与此相关的生命活动停止,引起机体迅速死亡。
b.解偶联剂 有些物质并不影响呼吸链中的电子传递,而解除氧化磷酸化的偶联作用,常将这类物质叫做解偶联剂。如2,4-二硝基苯酚(DNP)并不影响呼吸链氧化作用的正常进行,而使ADP不能磷酸化形成ATP。感冒或患某些传染性疾病时体温升高就是因为细菌或病毒产生的某种解偶联剂,影响氧化磷酸化作用的正常进行,导致较多能量转变成热能的结果。
c.离子载体抑制剂 有些物质可与K+或Na+形成脂溶性复合物,将线粒体内的K+转移到胞液。这种转移过程消耗了生物氧化过程释放的能量,从而抑制了ADP+Pi生成ATP的磷酸化作用。这些抑制剂有寡霉素、缬霉素、短杆菌肽等。
(2)胞液中NADH的氧化磷酸化 在糖代谢中糖酵解作用是在胞浆中进行的。在真核生物胞液中,NADH不能通过正常的线粒体内膜,要使糖酵解所产生的NADH进入呼吸链氧化生成ATP,需经过复杂的机制,现有的研究表明是通过α-磷酸甘油穿梭和苹果酸穿梭机制。
α-磷酸甘油穿梭作用主要存在于脑和骨骼肌中,苹果酸穿梭作用主要存在于肝和心肌中,如图7-5所示。
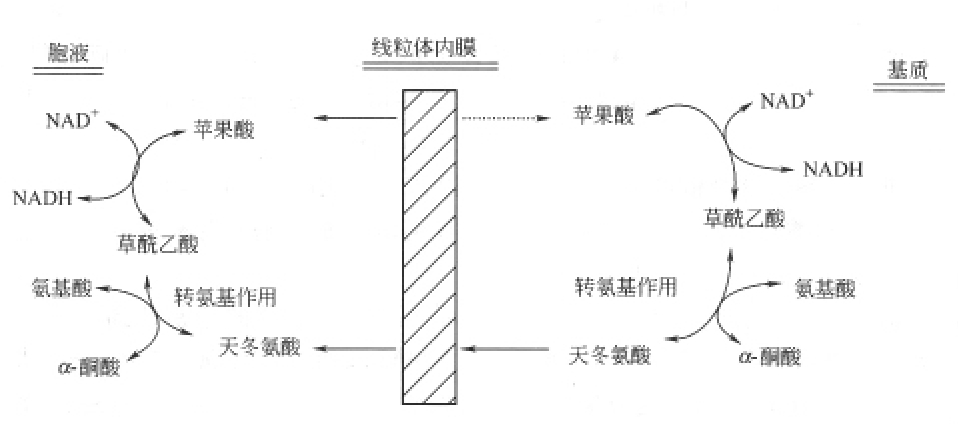
图7-5 苹果酸穿梭作用
五、非线粒体氧化体系
线粒体以外的氧化体系被概括为非线粒体氧化体系。非线粒体氧化体系主要包括发生在光滑内质网中的微粒体氧化体系和存在于微体中的过氧化体氧化体系。该体系与能量生成无关,其主要生理意义在于处理和消除环境污染物、化学致癌物、药物和毒物以及体内代谢有害物质等。
1.微粒体氧化体系
微粒体氧化体系存在于细胞的光滑内质网上,其组成成分复杂,目前尚不完全清楚。根据催化底物加氧反应情况不同,可将它们分为两种类型。
(1)加单氧酶系 加单氧酶系是由NADPH-细胞色素P450还原酶、细胞色素P450、FAD等组成的一种复杂酶系。其催化作用使氧分子中的一个氧原子被加到底物分子上,而另一氧原子与NADPH+H+上的两个质子化合成水。因催化作用具有双重功能又常叫做混合功能氧化酶,又因所催化的底物发生了羟化反应,还常称之为羟化酶。所催化的反应可简示为:
RH+NADPH+H++O2─→ROH+NADP++H2O
加单氧酶的主要功能在于:
①参与生物体内正常物质代谢。如肾上腺皮质类固醇的羟化、类固醇激素的合成、维生素D3的羟化以及胆汁酸、胆色素的形成等反应都与其有关。
②参与某些毒物(如苯并芘、苯胺等)和药物(如氨基吡啉、吗啡和苄甲苯丙胺等)的解毒转化和代谢清除反应。
(2)加双氧酶系 加双氧酶又叫转氧酶。催化两个氧原子直接加到底物分子特定的双键上,使该底物分子分解成两部分。其催化的反应通式可表示为:
R=R'+O2─→R=O+R'=O
例如,胡萝卜素加双氧酶催化β-胡萝卜素分解的反应如下:
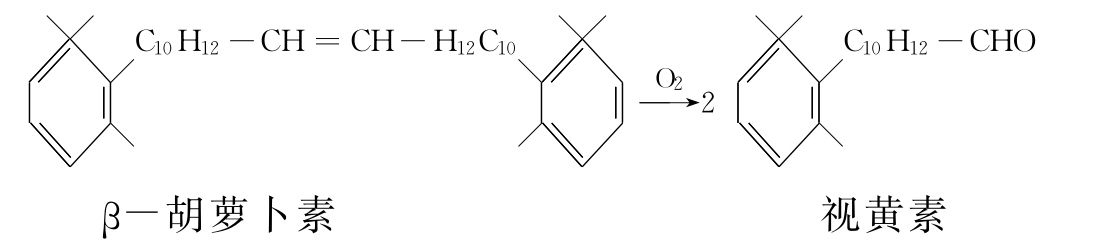
2.过氧化体氧化体系
(1)过氧化氢的生成 过氧化体中含有较多的需氧脱氢酶,它们可分别催化L-氨基酸、D-氨基酸、黄嘌呤等物质脱氢氧化,产生过氧化氢。
由于多种需氧脱氢酶的催化作用使机体产生了大量的过氧化氢,而造成对机体的影响。
(2)过氧化氢对机体的影响 过氧化氢对机体的作用具有两重性。有利的方面表现在:①在粒细胞和巨噬细胞中可杀死吞噬进来的有害细菌;②在甲状腺中参与酪氨酸的碘化反应有利于甲状腺素的合成等。然而,过氧化氢具有毒性,产生过多会对机体造成危害:①氧化含巯基的酶或蛋白质,导致它们丧失活性;②氧化生物膜中的不饱和脂肪酸形成过氧化脂质,损伤膜功能。过氧化脂质与蛋白质结合后进人溶酶体,难以分解排出,累积成脂褐素颗粒。
(3)机体对过氧化氢的处理和利用 在过氧化体中存在着能有效分解过氧化氢的酶类,它们可将过氧化氢转化为对机体无害的物质重新利用起来。
①过氧化氢酶 过氧化氢酶又称触酶,是含铁卧啉的结合酶,能催化过氧化氢分解为水和氧:
H2O2+H2O2─→2H2O+O2
②过氧化物酶 过氧化物酶也是含铁卟啉的结合酶。它可催化酚类或胺类物质脱氢,
并使脱下的氢与H2O2反应生成H2O。反应通式如下:
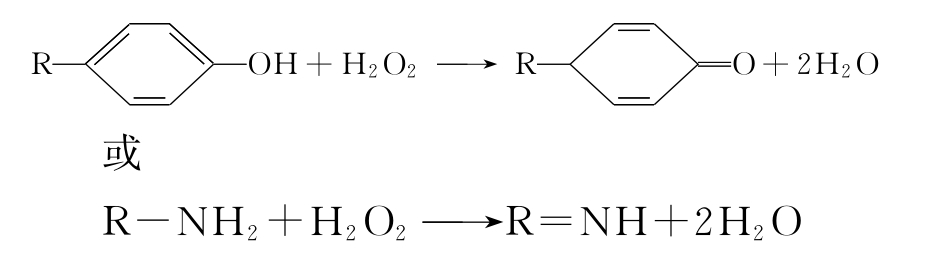
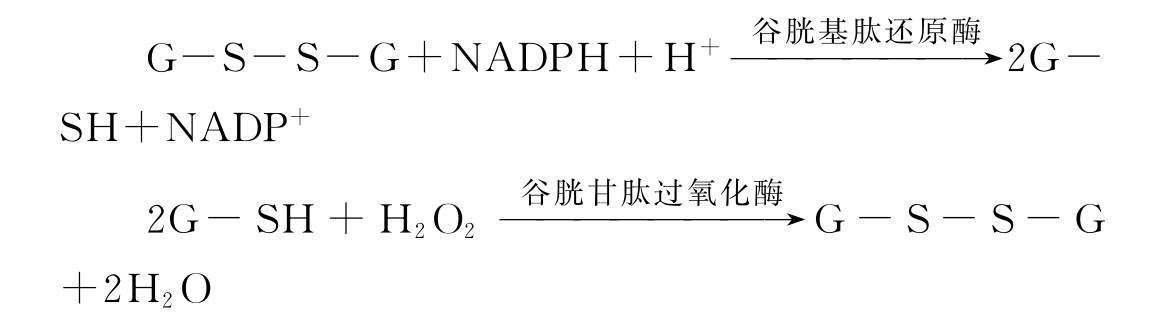
③谷胱甘肽过氧化物酶 在许多组织细胞中(尤其在红细胞中)存在着含硒的谷胱甘肽过氧化物酶。可催化还原型谷胱甘肽(G-SH)与过氧化氢反应,使过氧化氢分解,从而保护膜脂和血红蛋白免受氧化,维持它们的正常功能。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。










