



第四节 抗体生成的理论(Rules of Antibody Production)
抗体是由B细胞产生的。B细胞活化、增殖后可进一步分化为浆细胞,浆细胞可以在脾脏的红髓和淋巴结的髓质的生发中心,也可以移行到骨髓,成为抗原免疫后2~3周的抗体主要来源,骨髓的浆细胞可以在不依赖抗原存在的情况下,持续分泌抗体几个月甚至几年。抗体的合成与分泌与B细胞的增殖一样,需要CD40-CD40L介导的信号传递与细胞因子的作用,它们可以活化相关转录因子,增强Ig基因的转录和合成。
一、亲和力成熟的细胞分泌与抗原高亲和力结合的抗体
亲和力通常用来表示抗体对抗原结合力的大小,亲和力成熟是指在抗体生成过程中,凡是其BCR不能与抗原高亲和力结合者,均发生凋亡而被清除;极少数能通过FDC-B-T三种细胞相互作用,与抗原高亲和力结合的B细胞高表达bcl-2,则可进入下一轮增殖和突变。经历如此反复选择,最终存活的是表达高亲和力BCR的抗原特异性B细胞,其产生的抗体与抗原结合有较高的亲和力。亲和力成熟的主要原因是在生发中心有抗原刺激、有CD4+细胞辅助、有大量重组酶(RAG-1、RAG-2)的存在易发生以点突变为主要特点的体细胞高频突变,点突变经过积累效应,通过抗原选择逐步达到亲和力成熟(图12-8)。
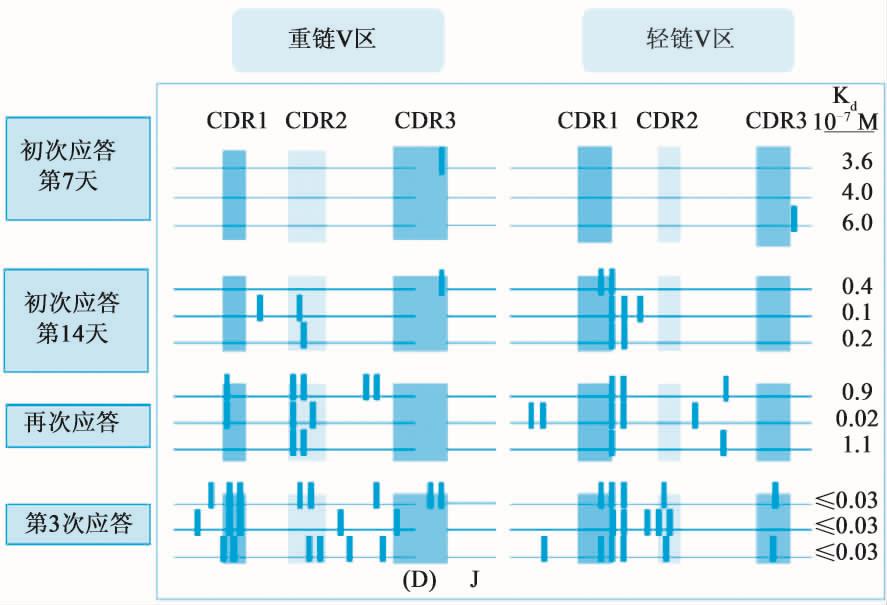
图12-8 B细胞亲和力成熟
二、Ig类别转换(Class switch)
经体细胞突变及亲和力选择而存活下来的B细胞可进一步发生Ig类别转换。Ig类别转换乃指抗体可变区不变(即结合抗原的特异性相同),但其重链类别(恒定区)发生改变。
1.影响抗体类别转换的因素
(1)细胞因子调节(图12-9)活化的T细胞分泌细胞因子是调控Ig类别转换的关键因素,如TH2细胞分泌IL-4促进抗体向IgE和IgG1转换,IL-5促进IgA产生;许多病毒、活化细菌Th1细胞、NK细胞分泌IFN-γ促进抗体向可以发挥调理作用、激活补体的IgG2和IgG3转换。
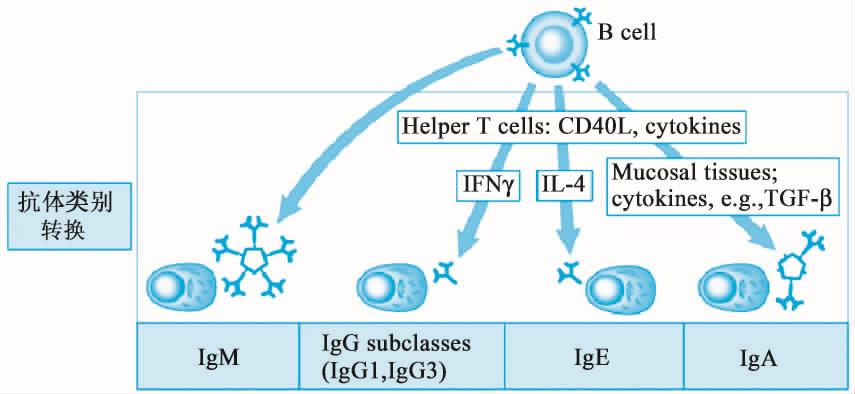
图12-9 细胞因子对Ig的调节
(2)抗原的性质TI抗原易诱导IgM产生,通常不引起抗体类别转换,但有些多糖对成年人还可诱导IgG2产生,对小鼠诱导IgG3产生;TD抗原主要诱导由IgM向其他Ig转换,其中可溶性蛋白主要诱导人和小鼠的IgG1;肠道寄生虫变应原主要诱导抗体向IgE转换。
(3)CD40L-CD40的作用CD40L与CD40可进一步激活转录因子NF-κB,诱导Ig类别转换。敲除CD40L与CD40的小鼠显示出Ig类别转换的严重受损,同时伴有亲和力成熟障碍和记忆性B细胞生成不良。
(4)免疫途径、免疫佐剂与转换的关系抗原免疫途径不同,转换类别也不同。皮内、皮下接种产生的抗体以IgG为主,而口服抗原则主要产生抗体IgA;抗原与Freund佐剂联合使用以产生IgG为主,而用铝佐剂则易诱导IgE产生。
(5)B细胞所在部位也与类别转换有关,特别是位于黏膜相关淋巴组织的B细胞主要向SIgA转换。
2.抗体类别转换的主要机制
抗体类别转换的主要机制是Ig恒定区基因重组或其重链mRNA的不同拼接即基因重排。
(1)Ig的基因结构 人重链基因定位于第14号染色体长臂,跨度约1 100kb,由多个不连续V、D、J和C四种基因片段组成(图12-10)。人VH基因的功能性片段约65个,编码重链可变区约98个氨基酸残基,包括CDR1和CDR2;人DH基因片段位于VH和JH基因簇之间,约27个,编码大部分CDR3;人JH基因位于DH下游,有9个JH基因片段(其中6个为功能性),负责连接VH和CH基因片段。JH编码约15~17个氨基酸残基,包括部分CDR3和FR4。人重链恒定区基因(CH基因)成簇排列,由多个外显子组成,排列顺序为5’-Cμ-Cδ-Cλ3-Cλ1-Cα1-Cλ2-Cλ4-Cε-Cα2-3’,跨度约200kb。每个外显子编码一个结构域,但铰链区由单独的外显子编码(表12-2)。人轻链基因分为κ和λ基因,分别定位于第22号染色体长臂和第2号染色体短臂。功能性V λ基因片段约30个,Vκ基因片段约40个。Vλ基因片段后是5个功能性Jκ和1个Cκ。每一个Ig其H链或L链基因分别由V、D、J、C基因片段群中各选择一个基因片段组合而成。
表12-2 人免疫球蛋白基因的染色体定位

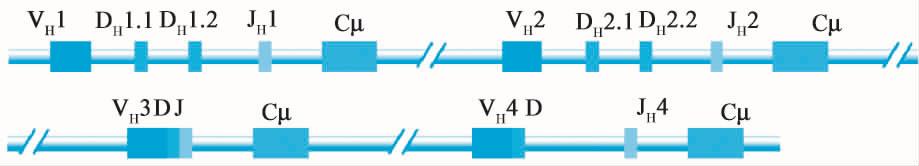
图12-10 免疫球蛋白重链和轻链胚系基因结构示意图
(2)基因重排 Ig恒定区基因重组或其重链mRNA的不同拼接即基因重排。由于Ig基因片段成簇存在,编码完整的功能性Ig肽链有赖于基因重排(gene rearrangement)。在编码C基因的5’端的内含子中,有一段含有[GAGCT]和[GGGGT]的重复性DNA序列称为转化区(switch region,S region),这种重复性序列在各Ig C基因有所不同,在类别转换为IgG3时,S和Cγ3发生重组,位于其间的Cμ,Cδ基因被环出,从而转录为γ3重链转录物。这种作用可以不止一次,可以转换到下游的其他基因,从而表达另一种Ig类别。即在V、D、J、C基因群中各选择一个基因片段,组成单个Ig的编码基因,进而转录和翻译,产生功能性Ig,此经体细胞突变及亲和力选择而存活下来的B细胞开始表达IgM,在免疫应答中首先分泌IgM,但随后即可表达和产生IgG、IgA或IgE,发生Ig类别转换。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。











