



第四节 T细胞的活化、增殖和分化 (T cells Activation,Proliferation and Differentiation)
一、T细胞活化的双信号理论
接受抗原刺激后,T细胞的完全活化有赖于双信号和细胞因子的作用。
1.T细胞活化剂
多种物质可活化T细胞:
(1)抗原(主要是TD-Ag);
(2)丝裂原(刀豆蛋白A,ConA、植物血凝素,PHA);
(3)抗T表面分子的mAb(抗CD3、抗TCRαβ、抗TCRγδ);
(4)超抗原(葡萄球菌肠毒素)。
2.T细胞活化的双信号刺激
此过程涉及T细胞与APC间的相互作用(图11-4)。
(1)T细胞活化的第一信号抗原识别信号即PMHC和TCR配接,提供抗原识别信号(第一信号)。APC将抗原肽-MHC分子复合物提呈给T细胞,TCR通过识别抗原肽形成TCR-抗原肽-MHC分子复合物。引起TCR共受体的交联,启动抗原识别信号,受体交联导致CD3、CD4(或CD8)分子的胞浆段尾部相聚,激活与胞浆段尾部相连的酪氨酸激酶,促使含酪氨酸的蛋白质磷酸化,启动激酶活化的级联反应,最终通过激活转录因子而导致细胞因子及其受体等的基因转录和产物的合成。
(2)T细胞活化的第二信号协同刺激信号(co-stimulatory signal)主要是B7和CD28配接,提供第二活化信号。仅有抗原识别信号尚不足以有效激活T细胞。APC和T细胞表面相应的黏附分子对(如B7/CD28、LFA-1/ICAM-1或ICAM-2/LFA-3等)结合,可向T细胞提供第二激活信号(即协同刺激信号),从而使T细胞完全活化。
CD28/B7是重要的协同刺激分子(co-stimulatory molecule,CM)。T细胞表达的CD28与专职APC上表达的B7-1(CD80)和B7-2(CD86)结合,可(经ITAM基序)发出(第二)活化信号。该信号的主要作用是促进IL-2基因转录和稳定IL-2 mRNA,从而明显促进IL-2合成。在缺乏协同刺激信号的情况下,IL-2合成受阻,抗原刺激不但不能激活特异性T细胞,反而导致T细胞失能(anergy)。
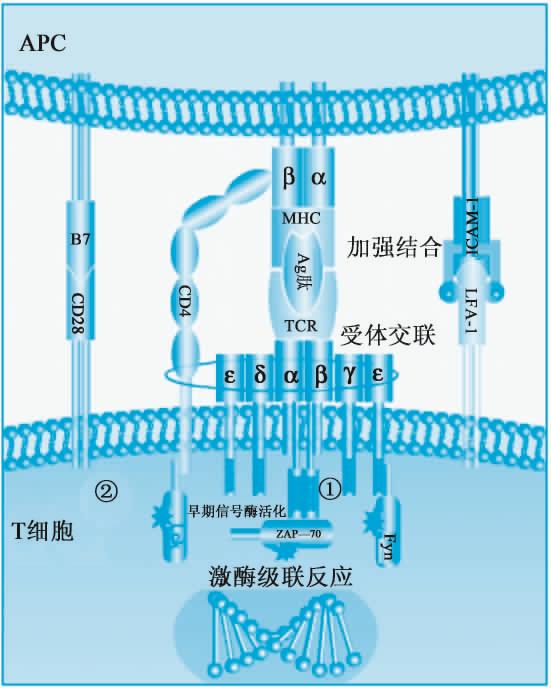
图11-4 T细胞与APC间的相互作用
表达CM的细胞只有几种特定的类型:如树突状细胞,单核Mφ,活化的B细胞等,这些细胞能高水平地表达强有力的CM,称为专职APC。
APC在静止状态CM的表达量很低,只有当病原微生物侵犯而有炎症时,炎症细胞释放CK,CK上调APC,APC才能表达CM。CM的表达或不表达,低表达或高表达,直接影响T细胞的应答。
此外,在T细胞激活至峰值后,还能表达另一种膜分子CTLA-4,CTLA-4与CD28具有高度同源性,该分子与B7的亲和力比CD28高约20倍。CTLA-4也能与B7-1/B7-2结合,但它的作用与CD28相反,可(经ITIM基序)向T细胞发出抑制信号,使活化的T细胞的子代细胞对抗原的敏感性降低,从而控制T细胞应答在一定范围。
根据T细胞活化的双信号理论,给予或不给予第二信号,可以人为地调节免疫应答,使之增强或抑制,为临床治疗提供新的途径。
除上述双信号外,T细胞的充分活化还有赖于细胞因子的参与。活化的APC和T细胞可分泌IL-1、IL-2、IL-6、IL-12等多种细胞因子,如IL-2和IL-2R配接,提供T细胞生长信号,它们在T细胞激活中发挥重要作用。
二、T细胞激活的信号转导
信号转导是免疫细胞激活的重要步骤。T细胞通过信号转导、T细胞识别信号被转换成胞内的生化事件,使信号进入细胞核,引起基因的转录激活和表达。
抗原和T细胞表面受体分子TCR的结合,引起细胞膜上各种跨膜分子如TCR/CD3、CD4(或CD8),CD45等相互靠拢成簇,使T细胞表面免疫突触的形成,胞内参与T细胞信号转导的蛋白酪氨酸激酶开始激活,引起一系列级联反应。
四条主要的信号转导途径参与T细胞的活化:①PLC-γ磷酸化启动的磷脂酯肌醇途径:下游分别通过PKC和钙调磷酸酶激活转录因子NF-ΚB和NF-AT;②MAPK相关途径:在Ras蛋白的参与下,通过MAPK中的Erk激活Jun和Fos,构成转录因子AP-1;③CD28介导的途径:通过PI3K促进转录因子NF-AF发挥作用;④IL-2R介导的途径:通过Jak1/ Jak3激酶等激活STAT5,并活化周期素E和Cdk2。
三、T细胞的增殖和分化
T细胞激活后产生以下结果:①细胞因子如IL-2等的合成与分泌,各种表面分子和受体如IL-2R的表达;②进入细胞周期,T细胞发生抗原特异性克隆扩增;③发生细胞因子依赖的功能性亚群分化;④记忆性细胞的产生和免疫记忆的形成。
双信号刺激只能诱导T细胞活化(这种活化的T细胞的数量是有限的),真正意义上的T细胞功能发挥,则依赖于扩增的子代效应T细胞。活化的T细胞分泌的多种CK,其中最主要的是IL-2,这些CK通过自分泌或旁分泌作用,使T细胞克隆扩增,并使子代细胞分化成效应T细胞。此外,IL-4、IL-12、IL-15等细胞因子也在T细胞的增殖和分化中(尤其是在Th1与Th2细胞的分化调控中)发挥重要作用。
T细胞经增殖(4~5天)后,分化为具有合成特殊效应分子能力的效应T细胞(Th细胞或CTL),发挥辅助、炎症、杀伤等免疫效应。同时,部分活化的T细胞可分化为长寿命记忆性T细胞,在再次免疫应答中起重要作用。
1.CD4+T细胞的增殖与分化
初始CD4+T细胞接受双信号刺激后,活化、增殖并分化为Th0细胞。Th0细胞具有分化为Th1或Th2细胞的潜能。Th0细胞的定向分化取决于局部微环境中细胞因子的调控,例如:IL-12可促进Th0细胞定向分化为Th1细胞;IL-4可促进Th0细胞分化为Th2细胞。Th0细胞的分化方向是决定机体免疫应答类型的重要因素。Th1细胞主要介导细胞免疫应答;Th2细胞主要介导体液免疫应答。
2.CD8+T细胞的增殖与分化
初始CD8+T细胞的激活主要有二种方式:
(1)APC直接提呈感染病毒的DC在提呈抗原肽-MHC分子复合物的同时,能高表达协同刺激分子,直接提供双信号刺激初始CD8+T细胞活化,使之合成IL-2,进而促使自身增殖并分化为效应T细胞。(图11-5A)
(2)Th细胞辅助由于CD8+T细胞作用的靶细胞(如肿瘤细胞、病毒感染细胞等)不表达或仅低表达协同刺激分子,此时,则依赖于APC及CD4+T细胞的协同作用才能激活初始CD8+T细胞。其机制为:病毒抗原、肿瘤抗原、同种异体MHC抗原从宿主细胞表面脱落后,以游离抗原形式被APC摄取,并在细胞内分别与MHCⅠ类或MHCⅡ类分子结合为复合物,表达于APC表面,分别提呈给CD4+和CD8+T细胞,引起两个细胞的活化。这种由一个APC细胞的双提呈,最终引起二个静止细胞的双活化现象,称为“交叉触发”现象。
①在交叉触发过程中,初始CD8+T细胞在APC-抗原肽-MHCⅠ类分子发出的特异性抗原识别信号的作用下,在初始CD4+T细胞接受APC双信号活化后分泌的IL-2的辅助下,增殖分化为效应T细胞(CTL)(图11-5B)。
②在交叉触发过程中,已活化的CD4+T细胞可促进APC表达B7等协同刺激分子,从而促使CD8+T细胞获得双信号而完全激活,并使之产生IL-2,引起自身增殖分化(图11-5C)。

图11-5 CD8+T细胞的增殖与分化
四、活化T细胞的转归
1.转化为记忆性T细胞,参与再次免疫应答
机体对特定抗原产生初次免疫应答后,部分活化的T细胞可终止分化转化为记忆性细胞(Tm)。当再次遇见相同抗原刺激时,Tm迅速增殖分化,产生强烈、持久的免疫应答。
2.适时发生凋亡,终止免疫应答
在免疫应答的后期,T细胞活化还可诱生一种重要的膜蛋白Fas及Fas配体(Fas ligand,FasL),二者结合后可启动Caspase酶联反应而导致细胞凋亡。
此外,在免疫应答的晚期,由于大量抗原被清除,T细胞所接受的抗原刺激和生存信号及所产生的细胞因子均减少,最终导致胞内线粒体释放细胞色素C,通过Caspase酶联反应而导致细胞凋亡。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。











