



元谋干热河谷苏门答腊金合欢、新银合欢人工林天然更新初步研究
马姜明1,2,李 昆1,张昌顺1
(1.中国林业科学研究院资源昆虫研究所,昆明650224;
2.中国林业科学研究院森林生态环境与保护研究所,北京100091)
摘 要:通过样地调查比较了苏门答腊金合欢、新银合欢的林分结实量、种子散布格局、种子库、及幼苗、幼树数量,并对影响天然更新的因素以及树种的适应性进行了初步分析;同时,使用灰色关联度法对两树种天然更新状况进行了综合评价。结果表明:①相同年龄新银合欢树种的单株结实量为1199粒·株-1,苏门答腊金合欢为566粒·株-1,同一树种单株平均结实量混交林高于纯林,天然更新的新银合欢林单株结实量介于新银合欢和苏门答腊金合欢之间;②随着距母树距离的增加,林地苏门答腊金合欢种子密度减少的幅度较新银合欢小,新银合欢种子的传播距离为90m,苏门答腊金合欢为110m;③苏门答腊金合欢人工林、新银合欢人工林及天然更新的新银合欢林关联系数分别为0.7269、0.6000、0.6000,苏门答腊金合欢天然更新效果稍好。
关键词:苏门答腊金合欢;新银合欢;天然更新;灰色关联度分析
1 引 言
森林天然更新是生态系统中森林资源再生产的一个重要生态学过程,是森林生态系统自我繁衍恢复的手段,同时也是森林群落动态的重要组成部分[18,24]。研究树种天然更新的特性以及影响天然更新的因素,对于了解森林生态系统的动态规律,采取合理的经营措施都是非常必要的。从目前的文献来看,对天然更新的研究主要是从树种的生物学特性、种子的产量和扩散、种子发芽、幼苗的生长情况及天然更新的影响因子等进行探讨[1,2,3,11,13,16,17,22,27]。
元谋干热河谷地处长江上游金沙江一级支流龙川江河谷下段。由于其特殊的气候条件,一直以来是植被恢复与重建的困难地区。近年来,在一批攻关项目的带动下,该地区在引种、造林树种的选择、树种抗旱性以及造林技术等方面进行了较为深入的研究,取得了较好的成绩,造林成活率明显提高。所引进的苏门答腊金合欢(Acacia glauca)和新银合欢(Leucaena leucacephala)这两个树种在干热河谷地区表现较好,种植面积逐年增加,目前在干热河谷地区的面积约7.0×103hm2。就元谋干热河谷地区的现状来看,能在该地区正常生长发育并能进行天然下种更新的树种在恢复与重建当地的生态环境显得非常重要,这样可以迅速地覆盖大片的裸地,达到恢复的目的。苏门答腊金合欢和新银合欢分属含羞草科金合欢属和银合欢属的灌木或乔木[7,9,29],两者均有速生、早实的特点,是目前干热河谷地区良好的造林树种。由于两树种的生物生态学特性不同,其天然更新的能力不尽相同。有关这两树种在该地区天然更新方面的研究还未见报道。探讨这两树种的天然更新对促进和维持元谋干热河谷地区人工林稳定性具有深远的意义。
2 研究地区与研究方法
2.1 研究地区概况
元谋县位于云南省滇中高原北部,金沙江中游一级支流龙川江下游。地理坐标为25°25'~26°07'N、101°35'~102°15'E;该区气候炎热,干湿季分明,年平均气温为21.9℃,最冷月(12月)月均温为14.9℃,最热月(5月)月均温为27.1℃,≥10℃的年积温为8003℃;年平均降雨量为630.7mm,雨季(5~10月)降雨量占年降雨量的90%以上;年均蒸发量为3426.3mm,为年均降雨量的5.4倍;年均相对湿度为55.8%,年均日照时数为2630.4h,无霜期为350~365d。其气候与热带稀树草原(Savanna)气候相似。
2.2 研究方法
2.2.1 标准地设置
本研究以20世纪90年代初所造的“长防林”为研究对象,天然更新的新银合欢林为附近的几株母树通过天然下种更新形成。在各试验地上选择具有代表性的地段,建立标准地,面积为20m×20m。各标准地的基本情况见表1。
2.2.2 母树结实量调查
采用标准株法。在各标准地内选取长势不同的母树10株,统计标准枝数量,选取3~5个标准枝,测定每个标准枝的平均结实量,求出标准株结实量,从而推算出林分总结实量。
2.2.3 林地种子数量调查
在种子脱落后期,在各标准地内设置5个1m×1m小样方,分别算出1m2内种子数,求出其平均数,推算各林分单位林地面积种子量。
2.2.4 单株母树种子散布距离调查
分别两树种选择与人工林同龄的孤立木3~5株,测得苏门答腊金合欢H=5.4 m,DBH= 3.6 cm;新银合欢H=5.8 m,DBH=4.1 cm。在距离母树每10m处调查种子密度。
2.2.5 林地种子幼苗和幼树调查
表1 各标准地林分状况及苏门答腊金合欢、新银合欢林分结实量(可靠性:95%,n=10)
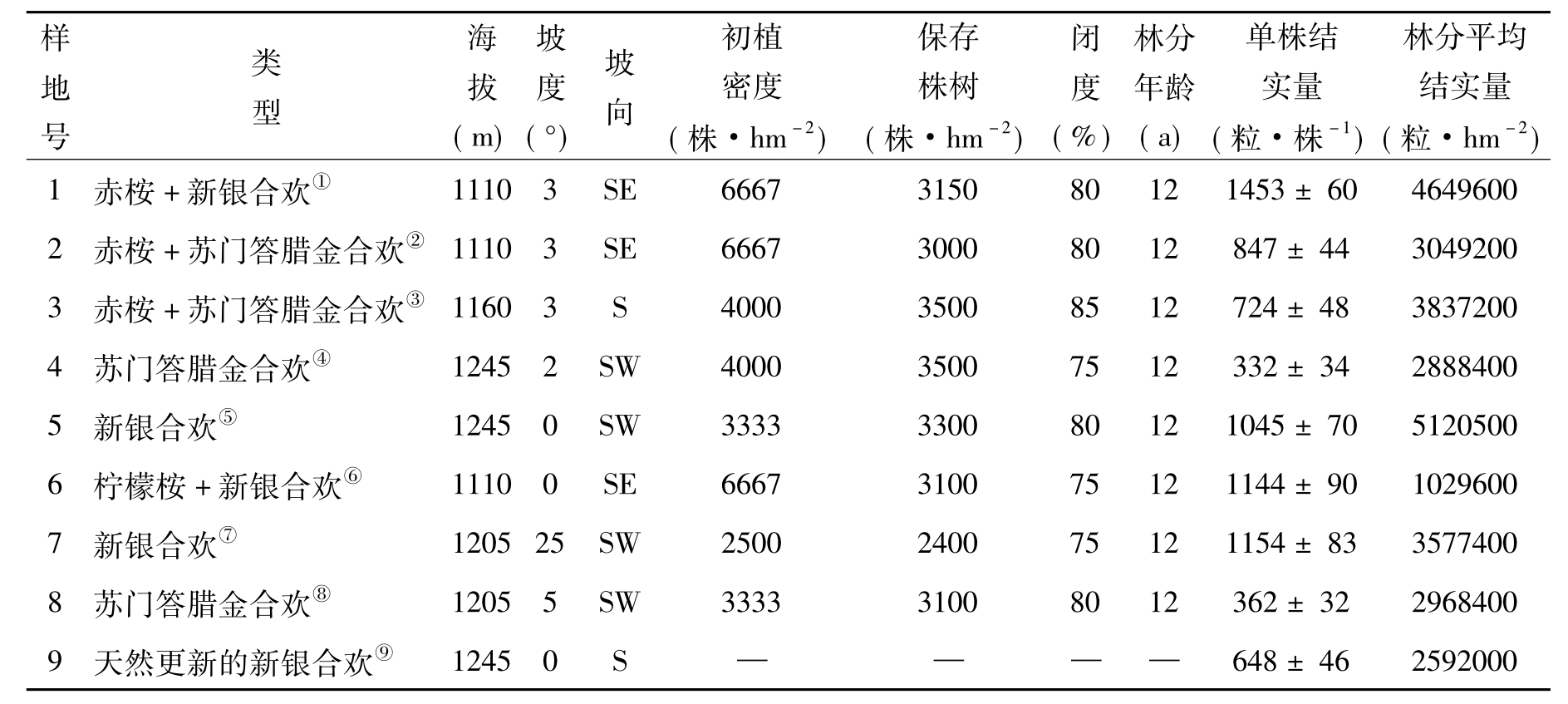
注:①Eucalyptus camaldulensis+L·leucacephala;②E.camaldulensis+A.glauca;③E.camaldulensis+ A.glauca;④A.glauca;⑤L.leucacephala;⑥E.citriodora+L.leucacephala;⑦L.leucacephala;⑧A.glauca;⑨Natural L.leucacephal.下同,The same below.
雨季初(2003年6月中旬)在已设立的样方内调查种子发芽数。由于研究地区土壤严重板结,分别把胚根裸露在土壤表面的种子发芽作为土壤表面发芽,而把胚根在土壤中的种子发芽作为土壤中发芽。在雨季中后期(2003年8月上旬)调查保存下来的幼苗数和调查样方内不同年龄幼苗或幼树的株数、高度、地径等。
2.2.6 用灰色关联度分析法综合评价树种天然更新状况
灰色关联度分析是灰色系统理论中因子分析及关系分析的主要方法。其原理是根据所研究的因子之间变化的相似程度来判断关联程度。能从多个因子来对天然更新状况进行综合评。具体方法为先对原始数据进行标准化处理,选择各因子达理想状态时的数值为参考点,组成参考数列,即x0=[1,1,…,1],运用公式△ij(p)=|xij-x0j|,i=1,2,…,n;n为各因子观察值个数;p=1,2,…,m; m为因子数,计算差值序列。然后求算△ij的最小值△min和最大值△max;根据公式Lij(p)=(△min+ 0.5△max)/(△ij+0.5△max)计算关联系数序列,根据Rij=∑Lij(p)/m计算关联度[5,30]。关联度越大说明因子间的变化态势越与理想状态接近,其相互关系越密切。具体在本研究中,评价天然更新的关联度越大,说明更新状况越好。本研究选择种子传播距离、1年生幼苗保存率及2年生、3年生、3年生以上幼苗或幼树数量等指标,综合评价苏门答腊金合欢人工林、新银合欢人工林、天然更新的新银合欢林的天然更新状况。
3 结果与分析
3.1 苏门答腊金合欢、新银合欢结实量、种子散布及种子库
从表1可以看出,新银合欢树种的结实量高于苏门答腊金合欢,新银合欢单株平均结实量为1199粒,苏门答腊金合欢为566粒,其单株结实量只有新银合欢的47%。同一树种新银合欢混交林中单株平均结实量为1299粒,新银合欢纯林单株平均结实量为1100粒。苏门答腊金合欢混交林中单株平均结实量为789粒,其纯林单株平均结实量为347粒,前者是后者的2.3倍。天然更新的新银合欢林单株结实量介于人工新银合欢和苏门答腊金合欢林之间。
种子散布和种子库是研究天然更新的重要组成部分。种子散布的研究可以了解种子成熟后的传播距离和散布格局等,这对掌握树种的更新特点非常重要。对种子库的研究可以得出种子的库存量,为种子的进一步萌芽提供物质保证。这两种树的种子主要以风播为主,其次为动物传播及人为传播。图1和图2分别为苏门答腊金合欢和新银合欢孤立木种子的传播距离和散布格局。可以看出,在距母树10m处,苏门答腊金合欢林地表面土壤种子库为107粒·m-2,低于新银合欢林地表面土壤种子库(189粒m-2),随着离母树距离的增加,两树种的林地表面种子密度逐渐减小。苏门答腊金合欢种子密度在距母树40m以后变幅有所减缓,而新银合欢在30m处。新银合欢种子的传播距离要比苏门答腊金合欢要短20m,苏门答腊金合欢种子的传播距离为110m。
3.2 苏门答腊金合欢、新银合欢更新
种子的萌发包括一系列的生理生化过程,是天然更新的第一步。只有在适宜的环境条件且种子的休眠特性不显著的情况下才能正常进行[26]。根据已有的研究,影响天然更新的环境条件主要是微环境、郁闭度、枯枝落叶层等[6,21,27,28],这最终还是归结到影响种子萌发和幼苗成长的温度、氧气和水分问题。此外,植物的化感作用、霉变等也影响到种子的正常萌发[4,12,15,19,20,23]。表2列出了不同苏门答腊金合欢、新银合欢林分天然更新情况。在6月中旬,1号、3号、5号样地每平方米种子发芽数较高,三者平均为112株·m-2,比总平均发芽数(87株·m-2)多出25株·m-2;4号、6号、7号样地发芽数较少,平均为65株·m-2,比总平均少22株·m-2。林地土壤表面发芽数普遍高于土壤中发芽数。这可能主要是土壤表面种子数量较多,种子在土壤剖面上具有递减的垂直分布[26],当年落下的种子绝大部分位于土壤表面,土层中的种子是以往年份落下的种子经过长期的雨水冲刷,动物活动和土壤裂缝的存在使得土壤逐渐向下移。至8月中旬再调查幼苗保存率时发现保存下来的幼苗较少,3号样地最多为5株·m-2,7号样地最少,只有1株·m-2。
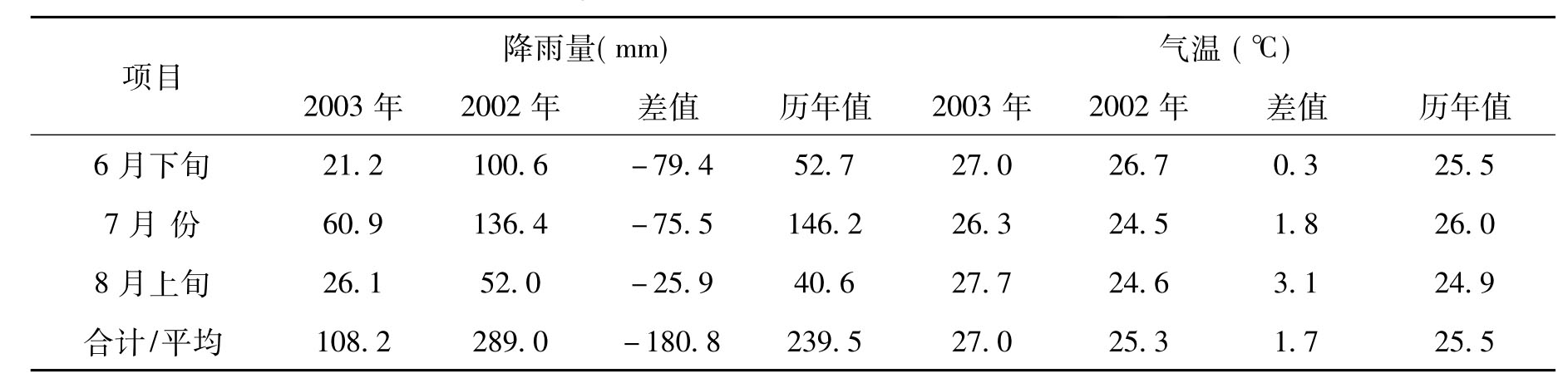
温度的波动会影响种子萌发,干季降雨量是幼苗成功定居的限制因子[8]。笔者对影响天然更新的气温和降雨量这两个主要因素进行分析,元谋属于干热河谷气候,干湿季非常明显。旱季从11月份开始到翌年的5月份,长达7个月之久。在此期间,降雨量仅占全年的10%,这两个树种的种子在雨季一到便立即萌发。2003年的雨季来得早,且5月份降雨相对充沛,这为种子萌发提供了很好的条件。但是自2003年6月以来该地区发生了较为严重的旱灾。从表3来看,6月下旬至8月上旬平均气温为27.0℃,与历年平均相比高1.5℃。再看降雨量,2003年6月下旬至8月上旬总计为108.2mm,是历年降雨量的45.2%。7月份降雨量为60.9mm,仅次于1976年(44.9mm)、1991年(57.4mm)的最少月雨量,比历年平均值偏少85.3mm。2003年而6月下旬至7月中旬这一时段降雨量为24.0mm;7月末至8月上旬降雨量仅为6.5 mm,这两个较长时间的干旱对幼苗的成活构成直接的威胁,从而导致8月份调查时成活的幼苗极少,尤其是土壤表面种子萌发的幼苗,难以从土壤中吸取所需的水分和养分,所以大多数1年生幼苗最终不能成活。相比之下,从土壤中长出的1年生幼苗情况稍好些。
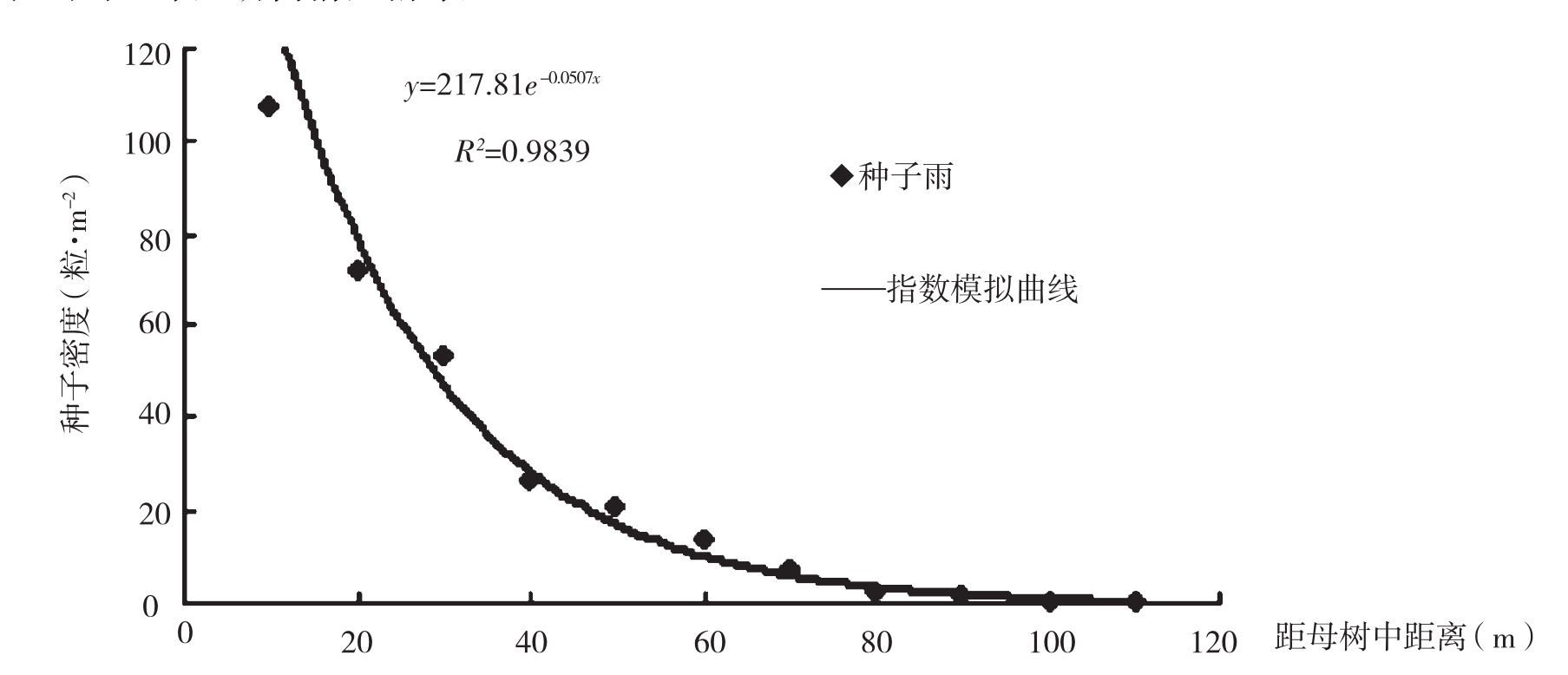
图1 苏门答腊金合欢孤立木种子雨
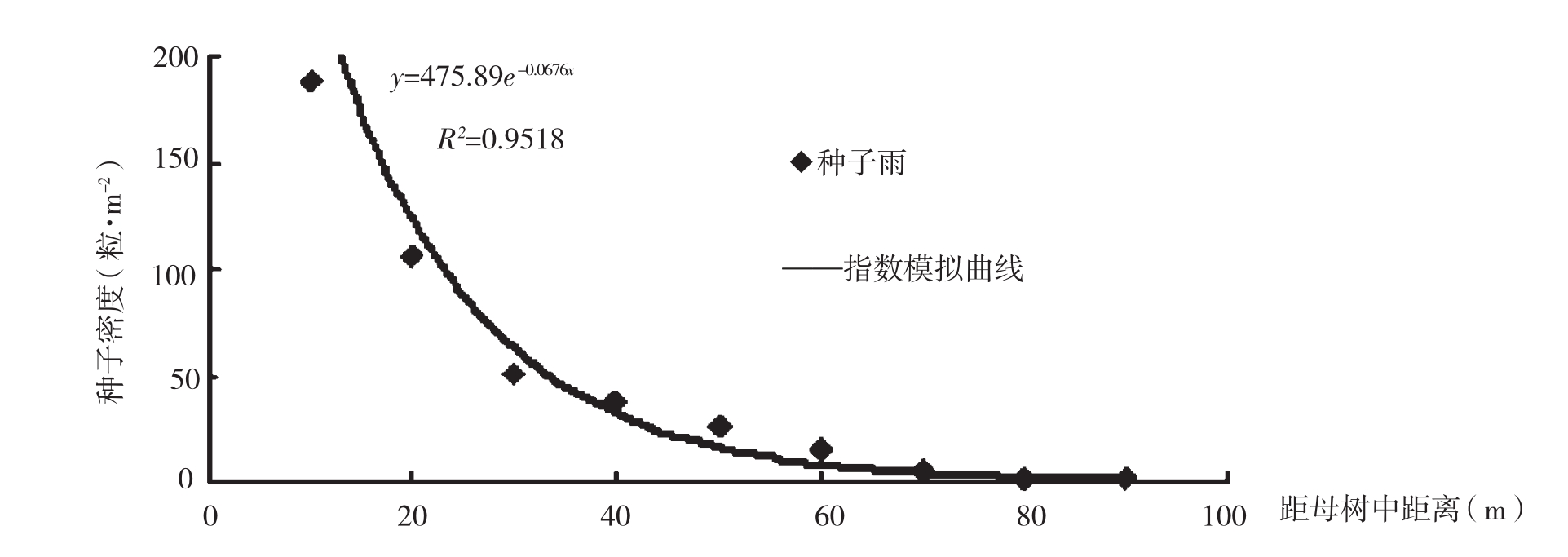
图2 新银合欢孤立木种子雨
再看各林分2年生以上的幼苗或幼树。从表2看出,不同林分2年生、3年生、3年生以上的幼苗或幼树变化较大,2年生幼苗或幼树平均为49株·m-2;最多为5号样地的新银合欢纯林,达93株·m-2,大部分集中在30~60株·m-2之间;最少的只有17株·m-2,为天然更新的新银合欢林。各林分3年生幼苗或幼树普遍比2年生少,平均为15株·m-2,只有2年生的35%。而3年以上幼苗或幼树更少,1号、2号、5号、6号、7号样地没有出现。3号、4号、9号样地在18~22株·m-2之间。这除了气候条件的影响外,还与幼苗生长发育过程中所需的营养物质有关,随着幼苗逐渐生长,需要的营养物质也就越多,幼苗之间争水、争肥现象突出,相当一部分幼苗得不到充足的养料而死亡,种群内受到密度制约的影响[14,25],出现“自疏”现象。调查中发现,3号样地赤桉、苏门答腊金合欢混交林天然更新较好,最高幼树为1.5 m,平均为0.9 m,这可能与该林地表面情况及人为活动相对较少有关。在侵蚀沟壑中或斜坡面等水分条件相对较好且光照条件相对适宜的微环境中,3年生以上的苏门答腊金合欢幼苗或幼树数量较多,在45~70株·m-2之间,高度在0.8~1.2 m之间,呈集群状分布,这也说明了微环境对天然更新的重要性。新银合欢在元谋的更新情况不太理想,除9号样地天然更新的新银合欢林外,其他样地没发现3年生以上新银合欢的幼苗或幼树。但是我们在元江干热河谷地区的公路旁、水沟边、崩塌地、冲积土、河滩等地调查发现,新银合欢能自然形成大片密集的异龄复层林,天然更新效果较好。通过调查该地区海拔360~680m高度,更新幼苗、幼树平均为45株·m-2,1年生幼苗为23株·m-2,2年生12株·m-2,3年生7株·m-2,3年以上4株·m-2。据胡琼梅研究[7],在元江县境内,海拔360~1500m范围的燥红壤、山地红壤上新银合欢均能繁茂生长。但在元谋的更新情况就不大一样,如果从气候上看,元江的年均气温比元谋高,年降雨量比元谋稍多,年蒸发量相对较少,空气相对湿度更大,这种气候条件似乎更适合新银合欢的天然更新,但目前有关更新机制的资料不多,有待进一步深入的研究。
表2 苏门答腊金合欢、新银合欢树种天然更新单位:粒·m-2,株·m-2
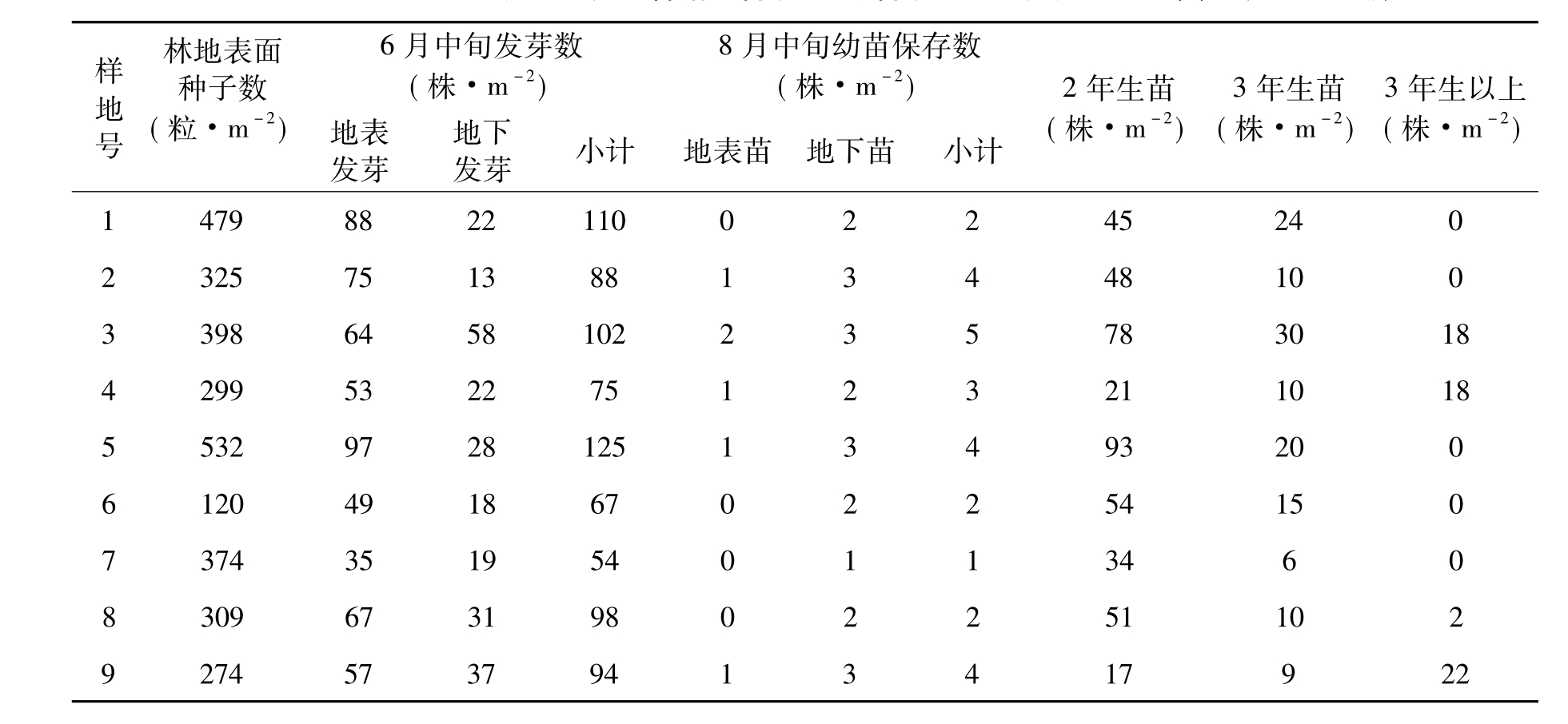
表3 2003年6月下旬至8月上旬元谋降雨量、气温变化
在元谋干热河谷地区所引进的众多树种中,苏门答腊金合欢、新银合欢这两个外来树种在天然更新方面显示出了它们的优势。从引种的气候相似性原理来分析,苏门答腊金合欢的原产地为加勒比海的库腊索岛(12°10'N,68°09'E),岛上年均气温为20.0℃,年均降雨量为580mm,属于热带半干旱气候,与元谋的气候条件相似[10]。新银合欢原产热带美洲地区,在年降雨量低于700mm的地区也能生长。这两树种都为耐干旱、耐瘠薄的阳生树种,在元谋干热河谷区生长良好,表现出较好的适应性。从生殖生态学方面考虑,苏门答腊金合欢、新银合欢与当地优势乡土树种如坡柳、山合欢、酸角等具有大致相同的结实物候期(见表4)。可以看出,这两种外来树种与当地乡土树种的开花期基本上出现在元谋的雨季,果实成熟期及种子脱落期都出现在旱季后期,也就是说恰好在雨季来临前,6月份进入雨季时,无休眠习性的成熟种子在适宜的气温和充足的水分条件下迅速萌发,实现由潜在种群向现实种群的转化。两外来树种1年生即可开花结实,早成熟的个体在幼体阶段停留的时间短,能够适应不良的外界环境,存活到繁殖的概率较大。这些都是苏门答腊金合欢、新银合欢树种在不稳定的环境中与其他物种竞争所形成的适应对策。
3.3 苏门答腊金合欢、新银合欢林天然更新综合评价
通过灰色关联度分析得出3种类型林分天然更新状况的关联度依次为:苏门答腊金合欢人工林(0.7269)、新银合欢人工林(0.6000)、天然更新的新银合欢林(0.6000)。关联度越大,说明天然更新状况越好。因此苏门答腊金合欢天然更新状况比新银合欢要好。从表5可以看出,苏门答腊金合欢各更新指标的关联系数分配相对比较均匀,而新银合欢各更新指标的关联系数分配落差较大。这说明苏门答腊金合欢在天然更新过程中的各个方面都表现出比新银合欢稍好的更新效果。用数学方法得出苏门答腊金合欢的更新效果比新银合欢要好,这与我们在调查时所发现的结果基本一致。
表4 苏门答腊金合欢、新银合欢及主要乡土树种在元谋的开花、结实物候期

表5 苏门答腊金合欢、新银合欢林分天然更新关联系数
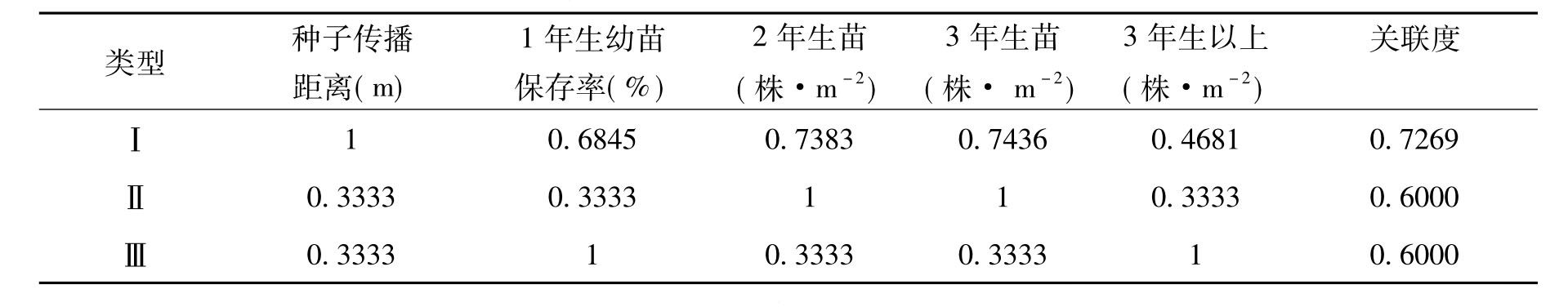
注:Ⅰ.苏门答腊金合欢人工林包括2号、3号、4号、8号样地;Ⅱ.新银合欢人工林包括1号、5号、6号、7号样地;Ⅲ.天然更新的新银合欢林,9号样地。
4 结论
(1)相同年龄新银合欢树种的结实量高于苏门答腊金合欢,同一树种单株平均结实量混交林高于纯林,天然更新的新银合欢林单株结实量介于新银合欢和苏门答腊金合欢之间。
(2)随着距母树距离的增加,林地苏门答腊金合欢种子密度减少的幅度较新银合欢小。新银合欢种子的传播距离要比苏门答腊金合欢要短。
(3)通过灰色关联度分析,苏门答腊金合欢人工林、新银合欢人工林及天然更新的新银合欢林关联系数分别为0.7269、0.6000、0.6000,说明苏门答腊金合欢天然更新状况比新银合欢林好。在侵蚀沟壑中或斜坡面等水分条件相对好且光照条件相对适宜的微环境中,3年生以上的苏门答腊金合欢幼苗或幼树数量较多。新银合欢更新效果在元谋不太理想,除了9号样地天然更新的新银合欢林以外,基本上没有发现3年生以上新银合欢幼苗或幼树。
(4)从本研究来看,今后应加强这两树种天然更新的机制研究,弄清更新过程的影响因素,进一步完善天然更新的评价指标体系及标准。
参考文献
[1]Allen J A.1997.Reforestation of bottomland hardwoods and the issue ofwoody species diversity.Restoration Ecology,5(2):125-134.
[2]Anderson L J,W interton A J.1996.Germination as a determinant of seedling distributions among natural substrates in Picea engelmannii(Pinaceae)and Abies lasiocarp(Pinaceae).Am J Bot,83(1):112-117.
[3]Anderson.L,Milberg P.1998.Variation in seed dormancy among mother p lants,populations and years of seed collection.Seed Science Research,8:29-38.
[4]Augspurger C,Kelly C K.1984.Pathogen mortality of tropical tree seedlings:experimental studies of the effects of dispersal distance,seeding density,and light conditions.Oecol,61:211-217.
[5]Deng J-L(邓聚龙).1987.Basal Method of Gray System.Wuhan:Huanzhong University of Science and Technology Press,(in Chinese).
[6]Gray A N,Spies T A.1997.M icrosite controls on tree seedling establishment in conifer forest canopy gaps.Ecology,78(8):2458-2473.
[7]Hu Q-M(胡琼梅).2002.Adaptability of Leucaena Leucocephala cv.salvador in Yuanjiang county of Yunnan province.Yunnan For Sci Technol(云南林业科技),3:61-65(in Chinese).
[8]Huang Z-L(黄忠良),Peng S-L(彭少麟),YiS(易俗).2001.Factors effecting seeding establishment inmonsoon evergreen broad-leaved forest.Journal of Tropical and Subtropical Botany(热带亚热带植物学报),9(2):123-128(in Chinese).
[9]Li J-Y(李纪元).2002.Resources of Australian Acacia and its potential of introduction to subtropical area of China.J Fujian For Coll(福建林学院学报),22(3):283-288(in Chinese).
[10]LiK(李昆).1998.Study on drought resistance and afforestation ofmain tree species in Dry-hotValley in Yuanmou county Yunnan province.Master dissertation,Southwest Forestry University,(in Chinese).
[11]LiX-X(李喜霞),Liu M-G(刘明国),Gao X-D(高晓东).2002.Factors of affecting natural regeneration of Chinese pine in gully of Chaoyang.J Shengyang Agr Univ(沈阳农业大学学报),33(5):352-355(in Chinese).
[12]Mallik A U.1995.Conversation of temperate forests into heaths:role of ecosystem disturbance and ericaceous plants.Environ Man,19:675-684.
[13]Meyer S E,A llen P S,Beckstead J.1997.Seed germination regulation in Bromus tetorum L.(Poaceae)and its ecological significance.Oikos,78:475-485.
[14]Nicotra A B,Chazdon R L,Iriarte S V B,et al.1999.Spatial heterogeneity of light and woody seeding regeneration in tropicalwet forests.Ecology,80(6):1908-1926.
[15]Nilsen E T,Walker J F,M iller O K,et al.1999.Inhibition of seeding survival under Rhodendron maximum L.(Ericaceae):could allelopathy be a cause?Am J Bot,86(11):1597-1605.
[16]Núnez M R,Calvo L.2000.Effect of high temperatures on seed germination of Pinus sylvestris and Pinus halepensis.For EcolMan,(131):183-190.
[17]Parciak W.2002.Seed size,number,and habitat of a fleshy-fruited plant:consequences for seedling establishment.Ecology,83:794-808.
[18]Peng S-J(彭闪江),Huang Z-L(黄忠良),Peng S-L(彭少麟),et al.2004.Factors influencing mortality of sees and seeding in plat nature regeneration process.Guihaia(广西植物),24(2):113-121(in Chinese).
[19]Rice E L.1984.A llelopathy(2nd ed).Orlando,Florida,USA,Academic Press.
[20]Sun S-C(孙书存),Chen L-Z(陈灵芝).2000.Seed demography of Quercus Liaotungensis in Donglingmountain region.Acta Phytoecol Sin(植物生态学报),24(2):215-221(in Chinese).
[21]Veblen T T.1989.Tree regeneration response to gaps along a transandean gradient.Ecology,70:541-543.
[22]Wang L-H(王林和),ZhangG-S(张国盛),Dong Z(董智).1998.A preliminary study on the seed yield and regeneration of Sabina vulgaris in Maowusu Sand.SciSilvae Sin(林业科学),34(6):105-112(in Chinese).
[23]Wu D-R(吴大荣),Wang B-S(王伯荪).2001.Seed and seedling ecology of the endangered Phoebe bournei(Lauraceae).Acta Ecol Sin(生态学报),21(11):1751-1760.
[24]Xu Z-B(徐振邦),DaiL-M(代力民),Chen J-Q(陈吉泉),et al.2001.Natural regeneration condition in Pinus koraiensis broad-leaved mixed forest.Acta Ecol Sin(生态学报),21(9):1413-1420(in Chinese).
[25]Yang Y-F(杨允菲),Zhu L(祝玲),Zhang H-Y(张宏一).1995.Analysis on the flux of soil seed bank and the seeding mortality in the communities of two species of Suaeda in the Songnen plain in China.Acta Ecol Sin(生态学报),15(1):66-71(in Chinese).
[26]Yang Y-J(杨跃军),Sun X-Y(孙向阳),Wang B-P(王保平).2001.Forest seed bank and natural regeneration.Chin J ApplEcol(应用生态学报),12(2):304-308(in Chinese).
[27]Zeng D-H(曾德慧),YouW-Z(尤文忠),Fan Z-P(范志平),etal.2002.Analysisofnatural regeneration barriers of Pinus sylvestris var.mongolica plantation on sandy land.Chin J Appl Ecol(应用生态学报),13(3):257-261(in Chinese).
[28]Zhang C-H(张慈海).1995.A preliminary study on natural regeneration of Pinus Massoniana.Sichuan For Sci Technol(四川林业科技),16(1):35-36(in Chinese).
[29]Zhang F-C(庄馥萃),Zhang X-Z(张雪珠),Chen Y-C(陈玉钗),etal.1993.Study on theadaptability and utilization ofAcacia glauca(L.)Moench.Plant introduction and acclimatization(植物引种驯化集刊),8:77-87(in Chinese).
[30]Zhang X-Y(张星耀).1999.Ecological and Mathematical Method of Forest Pathology.Beijing:Chinese Forestry Press,(in Chinese).
A preliminary study on regeneration of Acacia glauca,Leucaena leucacephala plantations inYuanmou Dry-hot Valley.
MA Jiangming1,2,LI Kun1,ZHANG Changshun1
(1.Research Institute ofResource Insects,Chinese Academy of Forestry,Kunming 650224,China; 2.Research Institute of ForestEcology,Environment and Protection,Chinese Academy of Forestry,Beijing 100091)
Abstract:An investigation was conducted on different plantations between Acacia glauca and Leucaena leucacephala about the natural regeneration from seed yield,the patterns of seed dispersal,seed pool and the number of seedlings and saplings.Preliminary analysiswasmade on factorswhich affectnatural regeneration,tree adaptability.Then synthetically evaluated the natural regeneration status of A.glauca and L.leucacephala with grey relational analysis.The result showed that the number of fructification of Leucaena leucacephala is 1199 grains per tree,of which A.glauca is 566 grains per tree at the same age.Whether A.glauca or L.leucacephala,themean number of fructification of individual in mixed forest wasmore than the one in pure forest.Themean number of fructification of individual natural regeneration L.leucacephala forest was between A.glauca and L.leucacephala plantations.As the distance to mother tree increased,the seed density of L.leucacephala on woodland decreased faster than that ofA.glauca.The seed dispersal distance of L.leucacephala 90m,of which A.glauca is110m.The grey relational coefficientofA.glauca plantation,L.leucacephala plantation and natural forest was 0.7269,0.6000,0.6000,respectively.The natural regeneration status of A.glauca plantation was better than the other two types.
Key words:Acacia glauca;Leucaena leucacephala;Natural regeneration;Grey relational analysis.
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。











