



空气中含有约80%的氮气,但植物、动物、人类和大多数微生物都不能直接利用这种气态氮作为氮素营养。自然界中只有极少一部分微生物可以将氮气逐步还原为氨而作为氮源。这种分子态氮的生物还原作用称为生物固氮作用。就世界范围来说,由生物固氮作用固定的氮素量要远比由工业固定的氮素量高。
(一)生物固氮机理
1.固氮反应及其基本条件
尽管能固氮的微生物多种多样,但它们固氮的基本反应都是相同的。
N2+8e+8H++nATP+Nase(固氮酶)→2NH3+H2+nADP+nPi
此反应很清楚地表明,要进行固氮必须满足以下基本条件:① 必须有具固氮活性的固氮酶。 ② 必须有电子和质子供体,每还原1分子N2需要6个电子和6个质子,另有2个质子和电子用于生成H2。为了传递电子和质子,还需有相应的电子传递链。 ③ 必须有能量供给,由于N2分子具有键能很高的三价键(N≡N),打开它需要很大的能量。 ④ 有严格的无氧环境或保护固氮酶的免氧失活机制,因为固氮酶对氧具有高度敏感性,遇氧即失活。 ⑤ 形成的氨必须及时转运或转化排除,否则会产生氨的反馈阻抑效应。
2.固氮酶的结构组成和催化特征
(1)结构组成
尽管自然界中固氮微生物多种多样,但固氮微生物所含固氮酶的组成大致相似,都是由2个亚单位即组分ⅠMoFe蛋白、组分ⅡFe蛋白和一个辅因子(FeMoco)组成。目前已知自然界中存在有3套含有不同金属的固氮酶,即在环境中无Mo时可被V代替,如环境中Mo和V都不存在时可由Fe代替,这些含不同金属的固氮酶的氨基酸序列同源性也有明显差异,特别是FeFe蛋白固氮酶与MoFe蛋白固氮酶的氨基酸序列同源性差异更大。但是大多数固氮微生物所含的是MoFe蛋白固氮酶,而且以MoFe蛋白固氮酶的固氮效率为最高(见表10-3)。
来自不同固氮微生物固氮酶的2个亚基之间可以进行互补,组成的固氮酶仍然具有固氮活性,但这种活性比各自原始的固氮酶活性要低。
(2)催化特征
固氮酶除了能催化N2还原为NH3外,还可催化还原下列物质:催化C2H2还原为C2H4, 2H+还原为H2,催化N3还原为NH3和N2,催化N2O还原为N2和H2O,催化HCN还原为CH4、NH3、CH3NH2、[C2H4,C2H6],催化CH3CN还原为CH4,CH3NH2、[C2H4、C2H6, C3H6、C3H8]。可见固氮酶是一个十分活跃、基质谱相当广的酶。但在所有能催化的基质中,以催化N2还原为NH3的反应效率最高。
表10-3 不同固氮酶的特性比较
(3)固氮酶的固氮催化机理
固氮酶催化的生物化学机理如图10-12所示。由图可见,固氮过程中,由呼吸作用、发酵、光合作用过程中产生的电子和质子首先还原NAD或NADP成为NADH或NADPH,由还原态的NADH或NADPH还原Fd或Fld,再还原固氮酶组分Ⅱ即Fe蛋白,由还原态的Fe蛋白还原固氮酶组分Ⅰ即MoFe蛋白,还原态的MoFe蛋白还原N2和其他各种底物。
图10-12 固氮酶的催化机制(引自周德庆,2002)
固氮酶合成、催化和酶活性调控的分子生物学研究已经相当深入,并已取得了丰硕的成果。固氮酶合成的各个基因结构及其功能已大多清楚。固氮酶nif基因簇表达在有氧和高浓度有效氮素因素下的调控机理也已阐明。
(二)固氮微生物与固氮体系
1.固氮微生物与固氮体系
生物固氮作用是固氮微生物的一种特殊生理功能。具有生物固氮能力的微生物生理类群称统为固氮微生物。共生固氮微生物指只有与高等植物或其他生物共生时才能固氮或有效固氮的微生物。自生固氮微生物指在土壤中或培养基上独立生活时而不与植物共生即能固氮的微生物。
根据固氮微生物是否与其他生物一起构成固氮体系,可分为自生固氮体系和共生固氮体系两大类型。依据固氮微生物与不同生物构成的共生固氮体系,可将它们分为豆科植物与根瘤菌共生固氮体系、联合共生固氮体系、蓝细菌与红萍共生固氮体系、蓝细菌与某些真菌形成地衣的共生固氮体系和非豆科木本植物与放线菌等的共生固氮体系。自生固氮体系又可分为光能自生固氮和化能自生固氮两种类型。
研究者们运用化学分析法、同位素N15标记法和乙炔还原法等,发现和肯定了自然界中许多微生物的固氮能力。至今已确定的固氮微生物(包括细菌、放线菌和蓝细菌)已近50个属。与固氮微生物可共生固氮的豆科植物约有700个属,非豆科植物有13个属。而且已经证明尽管固氮微生物多种多样,但都是原核微生物,至今尚未发现任何自然的真核生物具有固氮能力。固氮微生物在好氧性、厌氧性和兼性厌氧性,化能营养型、光能营养型、异养型、自养型等各个微生物生理类群中都有广泛分布。
2.自生固氮微生物
自生固氮微生物的种类很多,它们除了能固氮这一共同特性外,形态和生理特性各不相同。
好氧性化能异养型固氮微生物在自然界中最为普遍,其主要代表是固氮菌群。
固氮菌属(Azotobacter)细胞个体较大,幼年细胞为杆状,随着生长可变为球状,单个或成对,有时许多细胞不规则地聚集在一起。革兰氏染色负反应,多数能运动,有周生鞭毛,不形成芽孢,但能形成厚壁的孢囊。固氮菌可分泌大量黏液形成荚膜。能形成色素,有些种类产生非水溶性黄、棕、褐或黑色素,有些种可产生水溶性黄绿色荧光色素。固氮菌的细胞形态见图10-13。
图10-13 固氮菌在平板上的菌落和细胞形态
(引自http://inst.bact.wisc.edu/inst/i...id%3D274)
固氮菌的营养条件与一般腐生性细菌相似,可利用许多简单碳水化合物、醇和有机酸作为良好碳源。一般每消耗1g碳水化合物可固氮10mg。固氮菌除能利用分子态氮外,还能利用铵盐和硝酸盐等无机氮化物。在含有化合态氮的培养基中生长时不固定分子态氮。生长时还需要磷、钾、钙、硫、镁等矿质养分。各种微量元素尤其是钼对于固氮菌的生长和固氮都具有促进作用。固氮菌自身可合成必需的维生素类物质。但在培养基中加入吲哚乙酸、维生素B1和维生素C等时,可促进固氮菌的生长并提高固氮效率。
固氮菌为好氧性微生物,生长的氧化还原电位最适范围在Eh240~300mV之间。它们利用极高的呼吸率氧化有机物,对固氮酶进行防氧保护。固氮菌生长的适宜温度为25~30℃ ,高于45℃即会死亡。对湿度要求较高。对酸性环境很敏感,最适宜pH为7.0~7.5,生长范围为5.8~8.5。固氮菌能合成多种对植物生长有一定的刺激作用的维生素类物质,如生物素、环己六醇、烟碱酸、泛酸、吡醇素和硫胺素等。
其他的自生固氮菌有固氮单胞菌(Azomonas)、拜氏固氮菌(Beijerinckia)和德氏固氮菌(Derxia)等。另外还有节杆菌(Arthrobacter)和黄杆菌(Xanthobacter)等。
专性厌氧固氮微生物主要是一些发酵型的梭状芽孢杆菌(Clostridium),如最早分离的巴斯德梭菌(C.pasteurianum)。巴斯德梭菌是较大的杆菌,单生或成对,周生鞭毛。芽孢位于细胞中部或偏端,形成芽孢后细胞膨大呈梭状。最适生长温度为25~30℃ ,生长pH范围为5.0~8.5。此菌的固氮效率较固氮菌低,每消耗1g葡萄糖可固定1.5~7mg氮素。
专性厌氧的硫酸盐还原细菌群中,有些属的种具有固氮作用。如脱硫脱硫弧菌(Desulfo-vibrio desulfuricans)、普通脱硫弧菌(D.vulgaris)和巨大脱硫弧菌(D.gigas),瘤胃脱硫肠状菌(Desulfotomaculum ruminis)和东方脱硫肠状菌(D.orientis)等。近年来,已证明严格厌氧的产甲烷细菌中有一些种如巴氏甲烷八叠球菌(Methanosacrina barkeri)227菌株等具有固氮活性。
兼性厌氧固氮微生物主要包括肠道杆菌科和芽孢杆菌科的一些属种,如欧文氏菌(Er-winia)、埃希氏菌(Escherichia)、克氏杆菌(Klebsiella)、柠檬酸细菌(Citrobacter)、肠杆菌(Enterobacter)、芽孢杆菌(Bacillus)等。芽孢杆菌中的多粘芽孢杆菌(B.polymyxa)、浸麻芽孢杆菌(B.macerans)和环状芽孢杆菌(B.circulans)都能进行兼厌氧性生长和固氮。
化能无机营养型类固氮微生物至今发现和证实的仅有一种,即硫杆菌属(Thiobacillus)中的氧化亚铁硫杆菌(Thio.ferroxidans)。这是一种能固氮的独特的营养类型。
光合型固氮微生物可以分为光合细菌和蓝细菌两大类群。光合细菌群广泛存在于海域和淡水环境,又可分为紫细菌、紫色非硫细菌和绿细菌等非产氧光合细菌。它们都是原核微生物的G-细菌,具有特征性的细菌叶绿素a,只含一个光系统,营不放氧光合作用。蓝细菌群即蓝绿藻中的许多属种能进行固氮作用,现已证明24个属的120多个种具有固氮能力,其中大多属于念珠藻科,少数属于胶须藻科和蓝珠藻科。从形态上可分为有异形胞的蓝细菌和无异形胞的固氮丝状蓝细菌如鱼腥藻属(Anabaena)、念珠藻属(Nostoc)、柱孢藻属(Cylindrospor-mum)、单歧藻属(Tolypothrix),它们在自然界中有广泛分布。
固氮蓝细菌固氮量仅次于豆科植物与根瘤菌的共生固氮系统,且生理过程与放氧型光合作用相偶联,进化上有特殊地位,在固氮的同时也放氢,是一个有希望利用的太阳能生物转换系统。
3.根瘤菌和豆科植物的共生固氮
根瘤菌和豆科植物的共生固氮作用是一种最具有实际经济意义的生物固氮类型。共生生物固氮机理、根瘤菌的形态生理、遗传进化、生态学和分子生物学等都得到了深入研究。
(1)根瘤菌的形态和生理特征
实验室条件下根瘤菌为(0.5~0.9)μm×(1.2~3.0)μm大小的杆菌,革兰氏染色阴性,与其他土壤杆菌相似。细胞单个或成对,常可见成群排列。幼龄时能运动,快生型根瘤菌具周生鞭毛,而慢生型根瘤菌具单生鞭毛。无芽孢。幼龄细胞染色均匀,但老龄菌体细胞由于积聚有不染色且折光性强的聚β-羟基丁酸颗粒而导致染色不均匀或呈环节状。细胞外可形成大量黏液物质。
与豆科植物营共生时,根瘤菌形态经历不同变化。在从土壤进入根内时为很小的杆菌。随着根瘤发育,进入根瘤细胞内的菌体逐渐膨大或分叉,成为梨形,棒槌形,T形或Y形,这些特殊形态的根瘤菌称为类菌体。在类菌体形成之前,即可能开始固氮,在类菌体充分成熟阶段,即进行旺盛的固氮过程。快生型根瘤菌的大类菌体具有固氮功能而失去繁殖能力,而含菌组织中另一类正常的小杆菌仍保持有繁殖能力,从根瘤中分离培养的根瘤菌实际上是这种小杆菌的后代。
在糖类-酵母膏平板培养基上的根瘤菌单菌落为圆形,直径0.1~0.5cm,边缘整齐,无色、白色或乳脂色,有光泽。在培养基上各种根瘤菌的生长速度不同,豌豆,三叶草和菜豆等快生型根瘤菌,接种2天后即可出现菌落,4~5天后已较大。而大豆、豇豆等慢生型根瘤菌,接种3~4天后才有生长,1周后菌落生长充分。
根瘤菌营化能异养型生活,利用有机物作为碳源和能源。快生型根瘤菌可利用的碳源很多,如葡萄糖、果糖、甘露醇、蔗糖、阿拉伯糖等多种糖类、多元醇和有机酸。但慢生型种类对于碳源的要求比较严格,仅利用少数糖类如葡萄糖、戊糖等。根瘤菌分解糖类不产气,有些种类可产生微量的酸。
根瘤菌可利用铵盐或硝酸盐为氮源,但仅有无机氮源时生长不良,大多数根瘤菌需植物性氮素物质如酵母汁、豆芽汁等才生长良好。非共生条件下根瘤菌的氮源以化合态氮为主,分子态氮不能作为主要氮源。很多氨基酸可促进根瘤菌生长。根瘤菌生长需要各种矿质营养,尤其对磷的要求较高,其他如镁、钙、铁、钼、钴等元素也是必不可少的。根瘤菌生长还需要有丰富的B族维生素。
根瘤菌严格好氧,但对氧的要求不高。根瘤菌生长的Eh在150~450mV。大多数根瘤菌的最适生长温度在25~30℃之间。生长的适宜pH值在6.5~7.5之间。
根瘤菌一般对广谱性抗生素敏感。农业上广泛使用的杀虫剂、杀菌剂和除草剂对于根瘤菌的存活、结瘤和固氮效应的影响比较复杂,反应不一。
(2)根瘤菌与豆科植物的共生结构———根瘤
根瘤形成过程可以分为根瘤菌感染、根瘤发生和根瘤发育三个阶段,见图10-14。豆科植物根系在土壤中发育,分泌类黄酮化合物,刺激相应的根瘤菌在其根际大量繁殖。根瘤菌通过对豆科植物特定的识别作用,吸附在根表。在根瘤菌的作用下,根毛细胞壁变软,发生卷曲,根瘤菌从变软的植物根尖细胞壁处进入根毛,逐渐形成一条明显的套状侵入线。根瘤菌在侵入线内不断繁殖,并使侵入线不断伸长。当侵入线推进到皮层的3~6层细胞时,处在前方靠近内皮层的细胞受到刺激,分裂成为根瘤的分生组织。随着分生组织的不断分裂,根瘤不断长大,不断长大的根瘤又分化为不同组织,同时形成维管束并与根的维管束相连,以保证营养物质的输送。
图10-14 根瘤菌和豆科植物共生时的根瘤形成
根瘤菌在侵入线壁逐渐解体时释放到细胞中,先聚集在寄主细胞的细胞质外围,并开始迅速大量繁殖,充满细胞,形成含菌细胞组织。随后含菌细胞形成泡囊,每个泡囊初含一个细菌,随着细菌的几次分裂,每个泡囊中可含有8个细胞。此时的根瘤菌细胞形态变异为多形态的类菌体。此时一般不再分裂,类菌体充满寄主细胞。随着含菌细胞的形成,类菌体的泡膜里出现红色的豆血红蛋白。这是根瘤成熟的显著特征,标志着此时开始固氮。
根瘤是豆科植物和根瘤菌共生形成的特殊形态,是根瘤菌固氮的场所。因此豆科植物和根瘤菌之间的共生固氮量与四个因子相关。 ① 根瘤的数目;② 每个根瘤中含菌组织的容积和类菌体数量;③ 含菌组织所持续的时间;④ 根瘤菌各个种的比固氮活性等。一个有效的根瘤应含有丰富的豆血红蛋白,能保证供给固氮所需的能量,能及时地同化和转运固氮产物,避免产物的反馈抑制。
豆血红蛋白(leghaemoglobin,Lb)在根瘤中起着调节氧的缓冲剂作用,它向类菌体提供低浓度高流量的氧气,这样既保证固氮酶不因高氧压而失活,又能充分保证类菌体进行氧化磷酸化所需的氧气供给,使固氮酶呈现较高的固氮活性。豆血红蛋白的合成是由不同的基因编码的,其血红素部分是由根瘤菌携带的基因决定的,而蛋白质部分是由寄主豆科植物的基因编码的。两部分由各自的基因表达合成后再行装配为有效的豆血红蛋白。
根瘤中根瘤菌的生存、代谢活动和固氮作用都需要有足够的能量供给。类菌体所需的能源物质由豆科植物的光合产物输送到根瘤的含菌组织,然后通过氧化磷酸化合成ATP。在豌豆植株中每同化100单位的碳有32单位的碳被转运到根瘤中。
氨是固氮作用的最初产物,它一形成就从类菌体中分泌出来,这些氨必须立即被同化并转运,否则会对固氮酶造成反馈抑制。同化氨的第一步是由谷氨酰胺合成酶作用合成谷氨酰胺:
NH3+谷氨酸+ATP 谷氨酰胺+ADP+Pi
谷氨酰胺+ADP+Pi
谷氨酰胺合成酶可在氨很低浓度时起作用,因而可有效地同化和转运氨。在有NAD(P)H2提供电子时,谷氨酰胺与α-酮戊二酸反应成2个谷氨酸:
α-酮戊二酸+谷氨酰胺+NAD(P)H2→2谷氨酸+NAD(P)+
在有高浓度氨并有NAD(P)H2提供电子时,可在谷氨酸脱氢酶(GDH)催化下,氨直接与α-酮戊二酸反应生成谷氨酸:
α-酮戊二酸+NH3+NAD(P)H2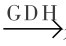 谷氨酸+H2O+NAD(P)+
谷氨酸+H2O+NAD(P)+
类菌体中每固定100单位的氨,只保留6单位的氨在根瘤中,绝大部分(91单位)转运给植株地上部分,而极小部分(3单位)转运给根系。
4.其他微生物的共生固氮
除豆科植物与根瘤菌之间的共生固氮作用外,还有内生菌和非豆科植物、蓝细菌和植物、蓝细菌和真菌之间的共生固氮以及细菌和水稻、玉米等禾本科植物的联合固氮作用。
(1)内生菌和非豆科植物的共生固氮
已知双子叶植物中能形成根瘤的有13个属的138个种,它们都是木本植物,如桤木(Al-nus)、杨梅(Mgrica)、木麻黄(Casuarina)、马桑(Coricuria)和沙棘(Hippophae)等,其中54个种已被证明结的根瘤具有固氮作用。它们根瘤的形成和结构与豆科植物根瘤不一样。在刚开始时,在根上出现小的突起,一两周后许多小突起簇生在一起,形成可达几厘米的根瘤簇,并可在根瘤簇的小球上又长出根来(图10-15)。根瘤内部结构与根的结构相类似,内生菌生存于皮层细胞中。
非豆科木本植物根瘤的共生菌是Frankia属放线菌(图10-15)。在形态上为纤细稀疏的菌丝体,菌丝分枝具横隔,发育后在菌丝顶端或菌丝间形成孢子囊。孢子囊为菌丝多向分裂形成的孢子堆构成,形态多样,大小不一。孢子囊内含有圆形或带角状、无鞭毛、不运动的孢子。Frankia放线菌能形成具有固氮功能的顶囊(vesicle),顶囊着生于顶囊柄上,再与菌丝相连。Frankia放线菌为微好气菌,有些种可产生黄、棕、褐等各种水溶性色素。利用Frankia放线菌接种试验表明,结瘤植物明显要比对照植株生长好,含氮量也高。
(2)蓝细菌和植物的共生固氮
红萍是热带和亚热带地区分布非常广泛、生长迅速、极为繁茂的水面蕨类植物。红萍是蓝细菌和蕨类植物的共生体。能固氮的红萍鱼腥藻(Anabaena azolla)生活于小叶鳞片腹部充满黏质的腹腔中,见图10-16。红萍提供光合作用产物作红萍鱼腥藻生活和固氮的碳源和能源,而红萍鱼腥藻则提供固氮产物。这种共生体的固氮效率很高,而且红萍含有很高量的易降解有机物,在我国南方是稻田中一种很有价值的水田绿肥。
图10-15 Frankia的菌丝(a)、顶囊(b)与形成的一种根瘤(c)
(引自web.uconn.edu/mcbstaff/.../Frankia/Frankia Home.htm)
图10-16 红萍与鱼腥藻的共生体剖面
(引自Francisco Carrapiço,SPIE Astrobiology Conference“Instruments,Methods,and Missions for Astrobiology IV”.San Diego,July29-30,2001.Proceedings of SPIE,4495:261-265,2002.ISBN 0-8194-4209-7)
蓝细菌念珠藻(Nostoc)或鱼腥藻(Anabaena)可以与裸子植物苏铁共生,使根形成反复二歧分枝形或珊瑚状,与正常根系完全不一样。用N15可证明这种珊瑚状根具有固氮作用。
在根乃拉草属(Gunnera)的植物叶片基部腺体中发现有念珠藻生存,并已用N15和乙炔还原法证明这些腺体中的念珠藻具有固氮活性。
(3)蓝细菌和真菌的共生
地衣是蓝细菌和真菌的共生体,在自然界中广泛分布于岩石、树皮、土壤,对于土壤的形成具有重要作用。在地衣中常见念珠藻、眉藻等,并已用N15和乙炔还原法证明具有固氮作用。固定的氮素可提供给真菌。
(4)联合固氮作用
联合固氮作用是指某些固氮微生物在植物根系中生活,并具有比在土壤中单独生活时高得多的固氮能力,但这种在植物根系中的生活方式又不同于根瘤菌和豆科植物根系之间的共生,两者既不形成共生体又较“松散”。在点状雀稗(Paspalum notaton)根的黏质鞘套内生存有一种固氮菌,定名为雀稗固氮菌(Azotobacter paspali),后又发现在热带牧草俯仰马唐(Digitaria decumbens)根系生活有固氮作用很强的含脂固氮螺菌(Azospirillum lipoferum) (图10-17)。随后在甘蔗、玉米、水稻等作物根际都测定到联合固氮活性。常见的联合固氮细菌有拜叶林克氏菌属(Beijerinckia)、雀稗固氮菌、固氮螺菌、粪产碱菌(Alcaligenes faecalis)、假单胞菌(Pseudomonas)和阴沟肠杆菌(Enterobacter cloa-cae)等。它们生活在根表的黏液区内,甚至进入根内细胞。
图10-17 Azospirillum lipoferum的细胞
(引自http://www.monografias.com/traba...iz.shtml)
(三)影响固氮效率的因素
1.氧对固氮酶的影响
固氮酶的一个重要特征是对氧具有高度敏感性,其催化固氮反应必须在无氧条件下进行。高浓度氧可对固氮酶的合成产生完全阻遏作用。分子生物学研究表明,随着氧浓度的变化,固氮酶的合成可被“启动”或“关闭”。氧对已合成的固氮酶两种组分可造成不可逆的损伤,使固氮酶丧失活性。而且氧可氧化固氮过程中的电子载体而使电子无法到达固氮酶。
好氧性固氮微生物的生长和固氮对氧的要求完全相反,这就要求具备保护固氮酶的防氧机制。实际上好氧性固氮微生物在长期进化过程中发展形成了多种防氧机制。这些机制中主要有加强呼吸强度的呼吸保护、利用构象改变来防止氧对固氮酶伤害的构象保护、具有较厚的荚膜或黏液层阻拦氧的渗入、固氮蓝细菌形成许多厚壁固氮异形胞和在根瘤含菌细胞中形成豆血红蛋白有效地为根瘤菌提供高流速低流量的氧用于氧化磷酸化,等等。
现已分离到一株其固氮酶对氧不敏感的嗜热自养链霉菌(Streptomycesthermoautotro-phicus),并发现该固氮酶的组成及其基因结构和固氮过程的电子传递都不同于原先的各种固氮微生物。
2.固氮作用中的氨效应
氨是固氮作用的产物,但氨的数量超过了固氮微生物机体本身的需要和迅速转换为氨基酸的能力时,积累的氨可阻遏体内固氮酶的生物合成。在缺乏NH4+的环境里,谷氨酰胺合成酶处于非腺苷化状态,具有催化功能和调节作用,能与固氮启动基因结合,推动RNA聚合酶催化转录mRNA,合成固氮酶。但在有丰富NH4+的环境中,谷氨酰胺合成酶被腺苷化,构象发生变化,失去与固氮酶启动基因区结合的能力,导致固氮酶不能合成。因此在培养固氮菌时如加入铵盐,则固氮菌不进行固氮而依赖铵盐生长。
3.ADP/ATP比率对固氮酶活性的调节
固氮酶催化N2为NH3,需要Mg2+和ATP参与,是固氮酶活性不可缺少的成分和正效应剂。Mg·ADP是Mg·ATP的水解产物,但其作用与Mg·ATP完全相反,是固氮酶的负效应剂,对固氮酶的底物还原活性部位起负的别构调节作用。它可以抑制从铁蛋白到钼铁蛋白的电子转移,并控制进入铁蛋白的电子总量,因而能有效地抑制固氮酶活性。因此细胞内ADP/ATP的比率可以调节固氮酶活性。有人认为ADP/ATP为1/2时,固氮酶活性可完全受到抑制。这种调节特性不仅与生物体的种类和生长条件有关,而且依赖于细胞内的Mg2+浓度。
4.环境中的C/N的影响
土壤中的C/N是影响固氮作用的最重要的因素之一。化能异养型固氮微生物只有在环境中有丰富的有机碳化合物而同时又缺少化合态氮时才能进行有效固氮。如果环境中化合态氮十分丰富,固氮微生物利用现成的氮化物作氮源,则固氮酶受到化合态氮的抑制,不显示固氮活性。另外,非固氮微生物由于氮源丰富而易于发展,与固氮微生物竞争碳源。因此只有在C/N比很高的环境中,这类化能异养型固氮微生物才会发挥固氮作用。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。













