



六、环境污染物的酶促降解
对进入环境的合成化合物的转化取决于微生物酶对这些非生物源化合物的认识和对稳定性结构(如卤代有机物的碳卤键)的催化反应。环境污染物的抗降解性(顽固性)部分是由于酶的活性和酶专一性的问题。降解酶对环境污染物的降解是十分重要的。卤代脂肪族化合物结构多样,应用广泛,如用作起泡剂(氯甲烷)、冷却液(氯乙烯)、土壤薰蒸剂(1,3-二氯丙烯、溴甲烷)、杀虫剂(六氯环己胺)。化学合成中的中间产物(1,2-二氯乙烷、氯乙烯、氯乙酸)和溶剂(三氯乙烷、三-和四-氯乙烯)。各种氯代烷烃化合物也见于生产过程的废弃物中。脱卤酶受到广泛而深入的研究,这里以脱卤酶为例来说明降解酶的专一性、反应动力学及酶的工程化改造。
1.脱卤酶酶催化脱卤
有学者把酶催化脱卤的方式归纳为氧化脱卤、脱卤化氢、取代脱卤、甲基转移脱卤和还原脱卤等五种。
(1)氧化脱卤(oxidative dehalogenation)
氧化脱卤在卤代脂(肪)烃和卤代芳香烃化合物的降解中十分重要,其脱卤过程是由单加氧酶和双加氧酶完成的,而其中有的反应是共代谢完成的,而有的反应则是以正常的代谢方式完成的。卤代烯烃、卤代短链烯烃和某些卤代芳香烃化合物可以通过共代谢方式实现酶催化降解(参阅后面的共代谢部分及图8-15),具有这种功能的是单加氧酶和双加氧酶。目前已知的有甲烷单加氧酶、氨单加氧酶,酚单加氧酶,2,4-二氯酚单加氧酶、甲苯单加氧酶、甲苯2,3-双加氧酶、丙烷单加氧酶、烯烃单加氧酶和异丙基苯双加氧酶。除了共代谢外许多微生物产生的单加氧酶、双加氧酶也能直接氧化卤代化合物脱卤。如假单胞菌CBS3菌株产生的4-氯苯乙酸,3,4-双加氧酶能氧进行氧化脱卤(图8-12A)。
(2)脱卤化氢
脱卤化氢酶从卤代有机底物消除HCl,导致形成双键。少动鞘氨醇单胞菌(Sphingomonas paucimobilis)UT26矿化农药γ-六氯环己烷(六六六)时会发生脱卤化氢(图8-12B)。
(3)取代脱卤(substitutive dehalogenation)
取代脱卤包括水解脱卤、硫解脱卤和分子内取代脱卤三种。
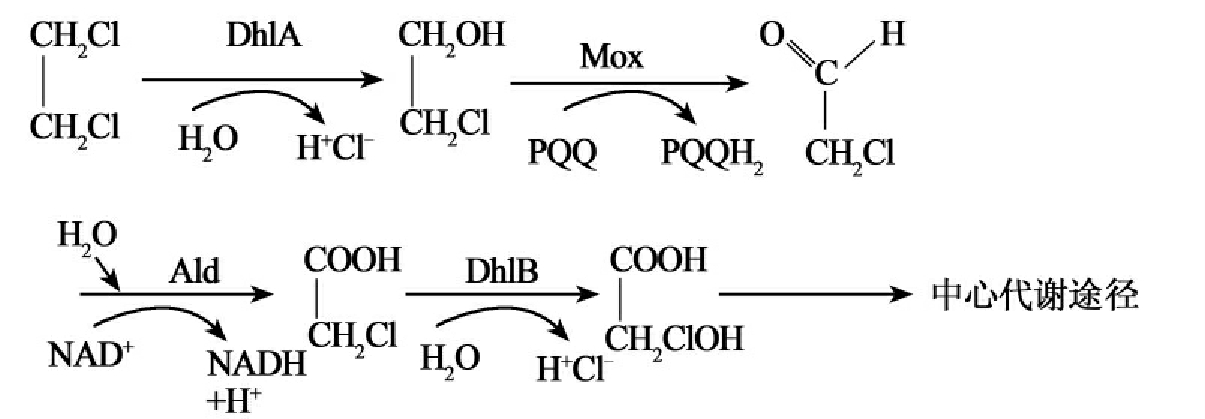
①水解脱卤(hydrolytic dehalogenation):被研究的脱卤酶大部分是水解脱卤酶。卤代的杂环类、芳香类和脂(肪)环化合物的水解脱氯已有许多研究报道。最先被分离纯化的卤代脂(肪)烃脱卤酶来源于自养黄色杆菌菌株GJ10。这种细菌可以利用1,2-二氯乙烷作为碳源(图8-12C)。
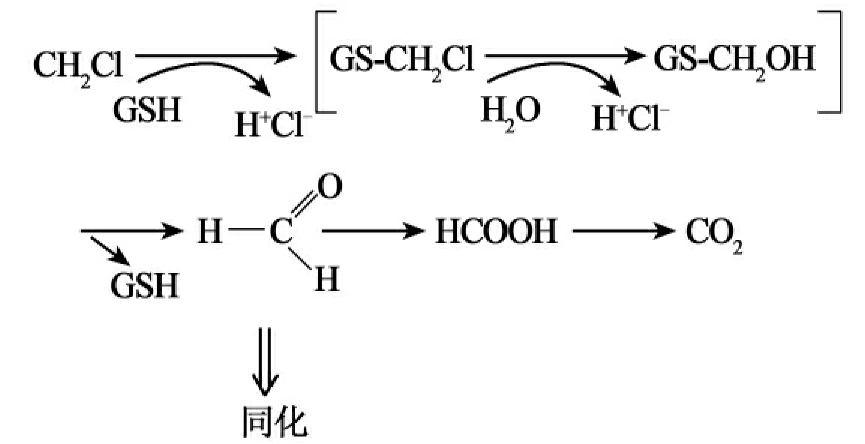
②硫解脱卤(thiolytic dehalogenation):谷胱甘肽S-转移酶催化形成氯化物和S-氯甲基谷胱甘肽。这种不稳定的中间产物被水解成谷胱甘肽,氯化物和甲醛,甲醛是甲烷营养菌生长的中心代谢物(图8-12D)。
③分子内取代脱氯(dehalogenation by intramolecular substitutive):卤代醇卤代氢裂合酶(Halohydrin hydrogen-halide lyases)(如卤代醇脱卤酶(haloalcohol dehalogenases)专一性催化邻位卤代醇和卤代酮的分子内取代反应,得到环氧化物。2,3-二氯-1-丙醇、1,3-二氯-2-丙醇、3-氯-1,2-丙二醇和其他卤代醇的降解(由假单胞菌菌株05-K-29、AD1等进行的)被认为是这种分子内的取代(图8-12E)。
(4)甲基转移脱氯(dehalogenation by methyl transfer)
甲基营养同型产乙酸(homoacetogen)脱卤醋酸杆菌(Acetobacteriamdehalogenans,即菌株MC)能利用氯甲烷作为唯一能源,产生乙酸。可被氯甲烷诱导的氯甲烷脱卤酶能转移作用底物甲基基团到四氢叶酸,产生甲基四氢叶酸和氯,甲基四氢叶酸被进一步代谢成乙酸盐,并通过乙酰-COA途径进一步反应(图8-12F)。
(5)还原脱氯(reductive dehalogenation)
还原脱氯对高氯代化合物(如PCDD和PCDF)的降解是十分关键的反应过程。
①共代谢还原脱氯(co-metabolic reductive dehalogenation)
许多产甲烷、产乙酸、硫酸盐还原和铁还原细菌能进行共代谢脱氯。这种共代谢主要见于卤代脂(肪)烃。反硝化假单胞菌菌株KC及腐败希瓦氏菌(Snewanella putrefaciens2000)菌株2000能通过共代谢降解三氯甲烷。这个过程受控于呼吸电子传递链的电子载体。细胞色素C、维生素K12被认为参与三氯甲烷的还原。
②耦联碳代谢还原脱氯(reductive dehalogenation linked to carbon metabolism):还原脱氯反应并不严格限于的厌氧细菌,紫色非硫细菌的深红红螺菌(R.rubrum)、度光红螺菌(R.photometrium)和血色红假单胞菌(Rhodospseudomonas rutila)在有氧条件下利用C2和C3卤代羧酸光能生长时,卤素取代基的还原是伴随着相应的羧酸的同化而进行的。
③呼吸过程中的还原脱卤(reductive dehalogenation as a respiratory process):许多菌株能把还原脱卤和能量代谢耦合在一起,它们以卤代脂(肪)烃或卤代芳香烃作为电子受体还原脱氯。一种硫还原细菌蒂氏脱硫念珠菌(Desulfomonile tiedjei)利用甲酸盐或H2作为电子供体,3-氯苯(甲)酸盐作为末端电子受体还原脱氯。还原脱氯和ATP合成的化学渗透假设的耦联(chemiosmotic coupling)已被证明。一种命名为Dehalococcides ethenogenes strain 195的细菌利用H2作为电子供体和PCE(五氯乙烷)为电子受体,完全脱氯PCE成为乙烯。PCE和TCE还原脱氯酶都是膜结合酶,并含有类咕啉辅因子。
A.
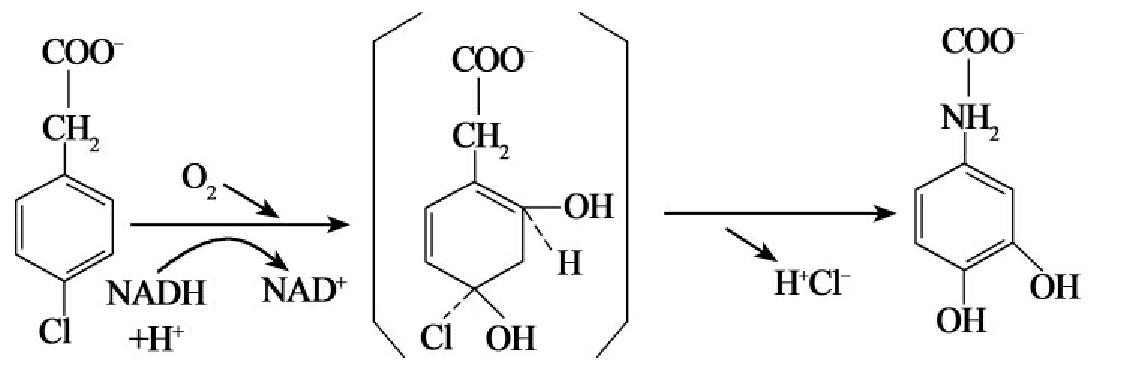
B.
C.
D.
E.
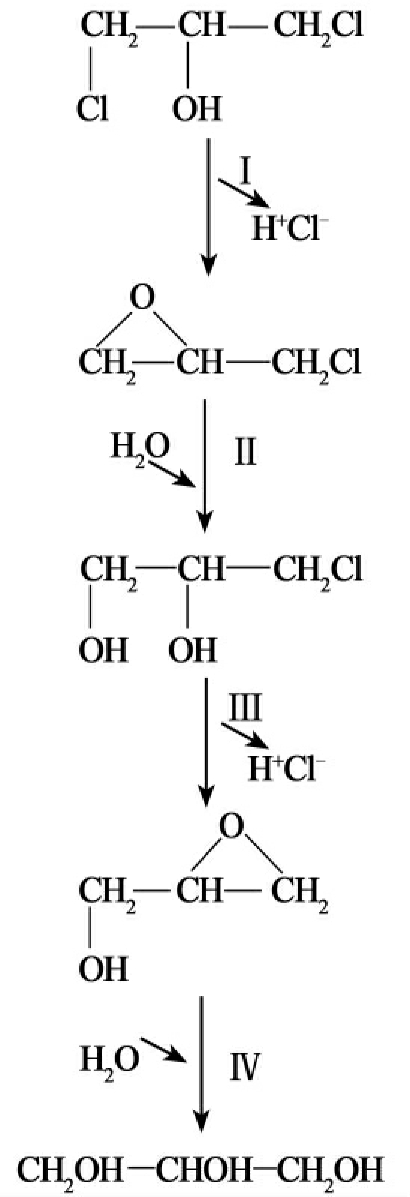
F.

图8-12 部分脱卤酶的脱卤反应
氯代烷烃脱卤酶催化C-卤键水解切开(cleavage)(图8-11C)的最先例子是自养黄色杆菌(Xanthobacter autotrophicus)和水生弯杆菌(Ancylobacter aquaticus)对1,2-二氯乙烷的降解,降解的第一步反应是由35kU的脱卤酶催化的,催化产生2-氯乙醇和氯化物。这种酶的底物范围很广,还能降解1,2-二溴乙烷,1,3-二氯丙烯以及长链氯烷烃等很多环境污染物。
卤代烷烃的降解酶中具有水解功能的是一组酶,一般属于α/β-水解酶,它们的活性中心中都有一个具有亲核攻击功能的ASP(天冬氨酸)残基。
自养黄色杆菌的脱卤酶(dehalogenase DhlA)的三维结构已被用x-射线晶体衍射法进行了分析。酶的活性中心是一个疏水的穴,位于具有α/β-水解酶折叠结构的球状主域(globular main domain)和分离的帽状区域之间。用x-射线晶体衍射法和同位素掺入法研究了催化原理。实验结果说明DhlA催化卤代化合物的断开是通过ASP124的羟化亲核取代(nucleophilic displacement)所进化的。产生的共价烷基-酶中间产物被活性水分子水解。水位于被脂化的ASP124的羰基碳上,HIS289以及ASP260通过减少一个质子使水分子活化。两种色氨酸包含在与底物卤素的结合中,面卤原子随后从底物中释放出来。
1,2-二氯乙烷是一种不存在于自然界的人工合成化合物,大量研究证明现在降解化合物的酶是近50年从一种更加原始的脱卤酶进化而来。支持其是最近进化而来的证据包括:①编码酶的基因的序列具有序列重复。②脱卤酶基因的组成型表达(黄色杆菌属和弯杆菌属菌株)。③基因定位质粒和从不同生物中分离到1,2-二氯乙烷脱卤酶缺乏进化趋异性。
2.酶的专一性
任何酶都有其底物专一性,酶的专一性分为两种类型,包括结构专一性和立体异构专一性。结构专一性按酶对底物专一程度的不同分为:绝对专一性(absolute specificity),酶对底物的要求非常严格,只作用于一种底物,不作用于任何其他物质。相对专一性(族专一性、基团专一性):酶作用底物不只是一种,对作用键两端的基团要求程度不同,对其中一端的要求严格,对另一个则要求不严格。键专一性酶只要求作用于一定的键,对键两端的基团并无严格的要求,这也是一种相对专一性,又称为“键专一性”,这类酶对底物结构的要求最低。立体异构专一性包括旋光异构专一性和几何异构专一性。旋光异构专一性是当底物具有旋光异构体时,酶只能作用于其中的一种。几何异构专一性是当底物具有顺、反两种结构时,只能催化其中一种结构,如延胡索酸水化酶只能催化延胡索酸即反-丁烯二酸水合成苹果酸,但不能催化顺-丁烯二酸的水合作用。
环境污染物的降解酶中有的是非专一性的,而有些有较强的专一性。共代谢转化是非专一性酶参与降解反应的重要例子,许多产生非专一性酶的微生物能降解非专一性的生理底物。如许多加氧酶不是专一性,如三氯乙烷可被发孢甲基弯菌(Methylosinus trichosporium) OB3b产生的溶解性甲烷单加氧酶(soluble methane monooxygenase)共代谢转化成一种环氧化物(epoxide),不稳定的环氧化物自发降解成各种产物。
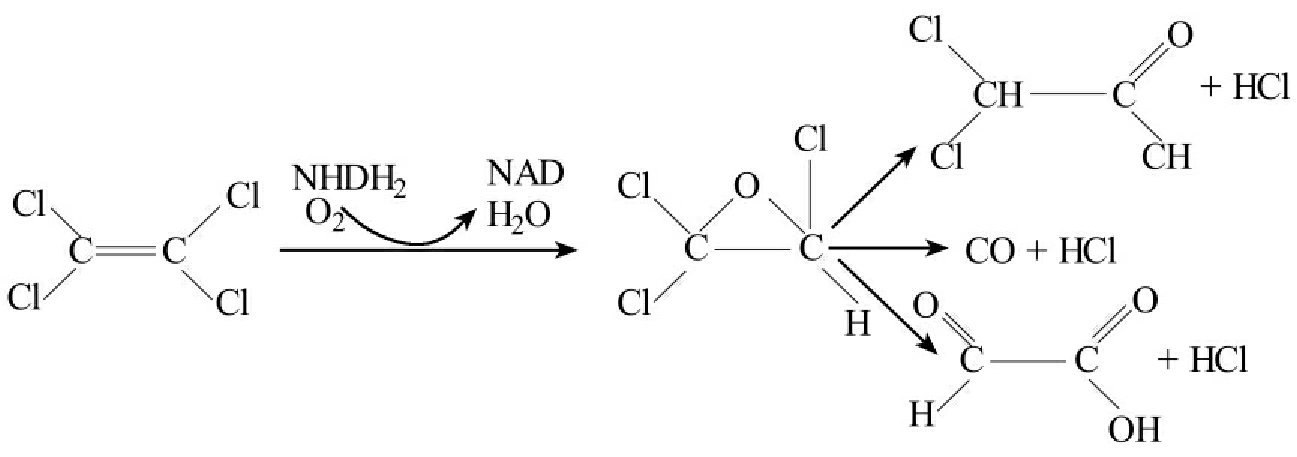
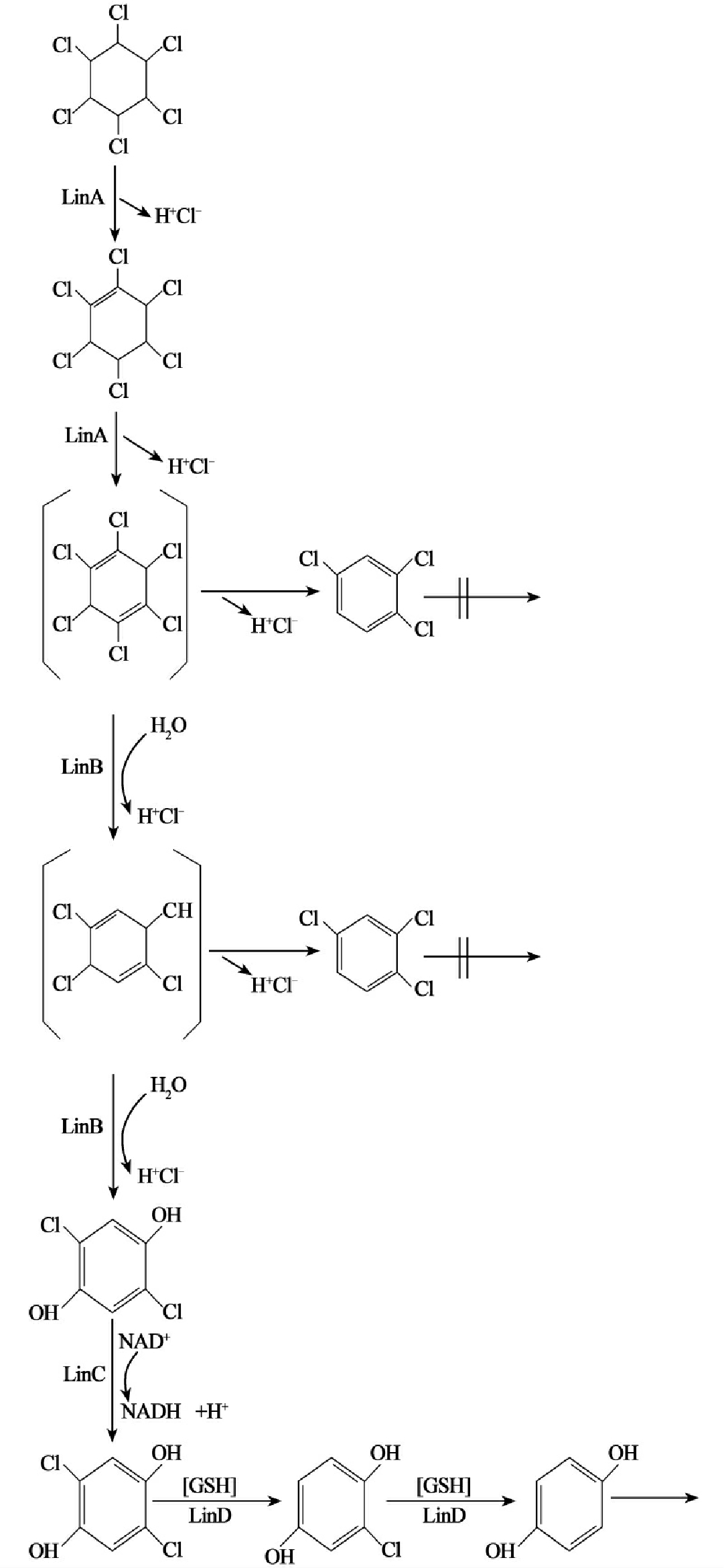
而利用卤代脂肪烃作为惟一碳源和能源的微生物在利用性降解时的降解酶是专一性的。碳卤键的断开可能取决于酶的专一性,但单个酶的底物专一性不一定和化学结构直接相关。在以氯代脂肪烃化合物作为碳源方面,化合物的结构特征是重要的,从这个意义上这类化合物的降解酶从总体上说有专一性,并具有一定规律性(图8-13)。
3.酶催化反应动力学
氯代烷烃的脱氯酶的反应动力学受到较为深入的研究。研究者用稳态(steady state)和前稳态(pre-steady-state)相结合的动力学实验确立动力学原理和推算与此相关的速率常数(图8-14)。
使用中止一流荧光淬灭实验(stopped flow fluorescence quenching experiments)证明酰化的卤素离子从包埋活性中心穴中的释放制约整个酶的转化速率,而酶的构象改变可使卤素快速释放,构象改变是酶的帽状区域(cap domain)的部分运动,这种运动使水进入到活性中心,并使卤素离子溶剂化。
对脱卤酶转化1,2-二溴乙烷和1,2-二氯乙烷所有步骤的速率都作了测定,发现C-Cl键的打开速率要比C-Br键慢得多,这使二溴乙烷的Km值比二氯乙烷小得多。卤化物的释放是1,2-二氯乙烷和1,2-二溴乙烷水解速度的主要限制步骤(表8-7)。对许多其他底物转化的动态动力学分析说明,酶对溴化合物的亲和力明显高于氯化合物,这是因为对碳-Br键有较高的打开速率和对溴化合物有较高的二级结合速率常数(higher second-order association rate constant)。一般来说对于相应易于转化的底物在高浓度情况下,溴和氯化同系物以同样的速率转化,这是因为卤素释放所需要的构象改变是速度限制因素。而对于难转化的底物,溴化的同系物的转化要比氯化的同系物快得多,这是因为碳—卤键的断开是速率限制的。
4.酶的工程化改造
(1)卤代烷烃降解酶突变体的筛选
大量的研究证明许多降解酶的突变体可以提高卤化物的降解和卤素释放速率。脱卤酶Val226 Ala突变体对1,2-二溴乙烷的转化的Kcat值高出野生型1.8倍。但Kcat的增加并不见于对1,2-二氯乙烷的转化(表8-7),这可以解释为碳-卤键断开速率的明显降低,造成这种结果的原因是C—Cl键断开速率明显低于C—Br键,对突变体来说碳—卤断开成为对1,2-二氯乙烷转化的最慢的步骤。从这里也可以看到提高转化1,2-二氯乙烷酶的活性比提高1,2-二溴乙烷的酶活性要困难得多。提高活性的效应也见于卤代烷烃脱卤酶phe172Trp突变体(表8-8)。Phe172是螺旋-环-螺旋结构覆盖的活性中心穴疏水帽状区域残留基中的一个。Phe172Trp突变体酶比野生型对1-氯乙烷有10倍高的Kcat/Km值。研究表明其活性中心螺旋-环-螺旋结构的可塑性比野生型酶更高。这种增加使它更易结合1-氯己烷这样的大底物,有更强的亲和力,而野生型的活性中心穴则不适合于这种基质。对1,2-二溴乙烷转化速率的增加是由于在卤素释放前酶的异构化速率(rate of the enzyme isomerzation)的提高,这与前面提到的Val226Ala突变体情况一样,而异构化可能实际上是帽状区域部分构象的改变。

图8-13 结构特征和微生物可以利用作为生长基质的卤代化合物的关系破折线左边化合物能支持一种或多种微生物的生长,而右边则为难以利用的同系化合物。
![]()
图8-14 代烷烃脱卤酶的动力学模式
1.形成米氏复合物;2.碳卤键断开;3.共价中间产物断开;4.构象改变和卤素脱出
表8-7野生型卤代烷烃卤酶和Val226Ala突变体(226Val Ala突变体)四步反应的动力学常数
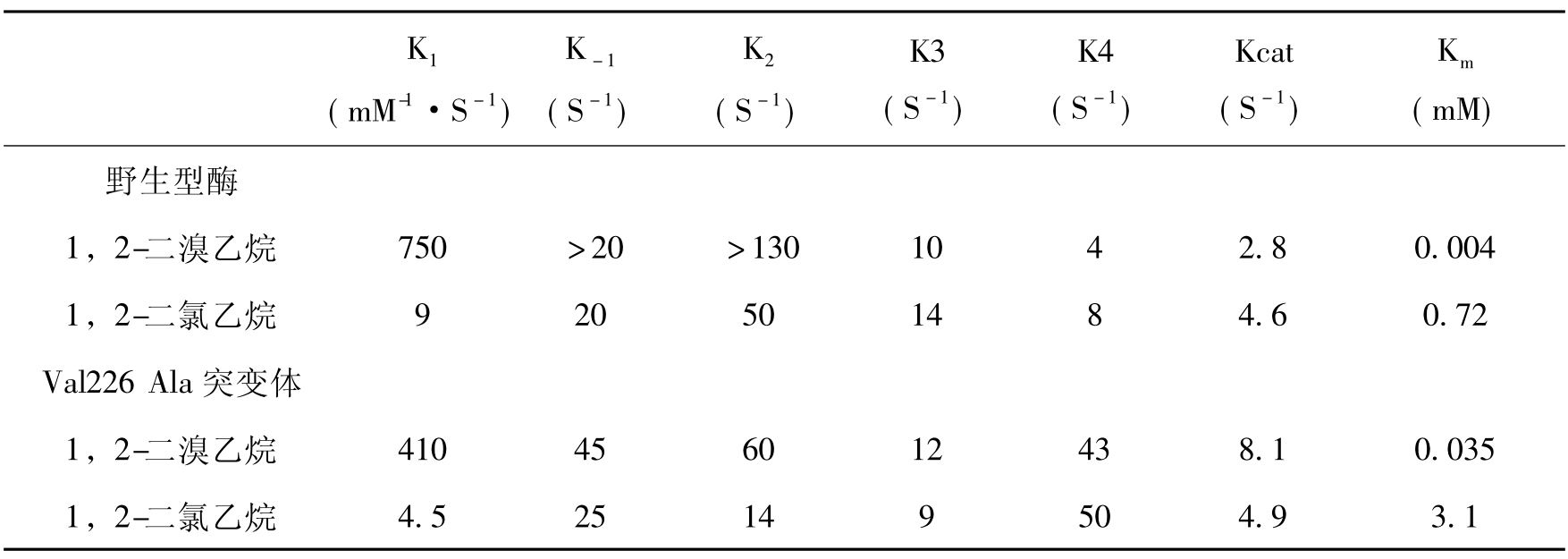
表8-8野生型、Phe172Trp(体外构建)和Asp170His(体内选择)卤代烷烃脱卤酶转化1,2-二氯乙烷、1,2-二溴乙烷和1-氯己烷的专一性常数(Kcat/Km值)

(2)新专一性的体内选择
通过对酶突变体的体内选择可以选择出降解原来不能降解底物的突变体,即为选择新的专一性,新专一性的体内选择。通过对自发突变的选择使脱卤酶适应新的底物,突变体能利用野生型酶难以水解的1-氯己烷。研究表明帽状区域的N端部分有短的定向重复,其对卤代烷烃脱卤酶适应新底物起重要作用。两个定向重复是15bp的优先重复和9bp重复。如果缺失两个重复就会失去1,2-二氯乙烷的水解活性,但仍具有对几种溴代脱卤酶底物的活性。这说明编码帽状的N端部分定向重复的DNA序列一定是最近进化出来的。而这个新的酶是在一个较老的脱卤酶是对工业产生的二氯乙烷适应中选择的。
(3)构建多酶体系的工程菌
对低分子卤代脂肪烃转化能力的瓶颈来源于断开碳-卤键和反应中间产物对生物的毒性。因此通过在合适生物中表达脱卤酶基因和构建带有多种能攻击顽固的结构和有毒的反应中间产物的酶的生物,可以提高微生物对化合物降解能力。有人已成功构建出基因工程菌,可以使极难降解的四氯化碳得到降解。
(4)利用已有的能转化非生物源化合物的有价值酶作为资源,开发具有区域和立体构型转化效应多样活性的酶。
(5)蛋白质工程的定位突变
定位突变技术是按照预先设计,精确地使靶基因在特定位点发生碱基序列的变化,进而使基因表达及其调控、基因产物发生相应改变。利用这种技术可以对目前的降解酶做进一步的工程改造,从而产生出具更强降解能力的酶。降解酶的定向改造和进化,见本章第一节。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。










