



7.1.3 Cu,Zn-SOD基因的克隆与表达
Cu,Zn-SOD基因已经从多种微生物、动物、植物等生物中克隆得到,并且有部分在大肠杆菌、酵母、病毒、昆虫、哺乳动物等生物体中得到表达。SOD基因的克隆主要通过以下几种方式完成。如利用E.coli sodAsodB双突变体菌株不能在氧化性培养基或基本培养基上生长的特点,通过功能互补筛选含有SOD基因的DNA片断,克隆SOD基因。根据SOD保守氨基酸序列设计探针,利用杂交的方法筛选含有SOD基因特征序列的DNA片断,进而克隆获得SOD基因。此外,PCR方法以及构建基因组文库克隆SOD基因的方法也有广泛应用。在大肠杆菌中,外源SOD基因表达量在菌体可溶性蛋白中占有相当高的比例,表达产物稳定性好,而且表达产物大多具有生物学活性,还有一些表达产物表达出对极端条件有很高的耐受能力。
7.1.3.1 人Cu,Zn-SOD基因的克隆与表达
A.人Cu,Zn-SOD基因在大肠杆菌中的克隆与表达
人源SOD作为一种重组蛋白由于在大肠杆菌中过度表达等原因,使工程菌合成的SOD蛋白质分子不能及时折叠成正确天然的二、三级空间结构,表达产物形成包含体,需要经过复性过程,才能转化为可溶性的有活性状态。
袁勤生实验室进行了人Cu,Zn-SOD基因在大肠杆菌中的克隆与表达,均获得了可溶性的表达,表达的重组Cu,Zn-SOD活性高。步骤如下:取构建好的pET-SOD分泌表达载体转化大肠杆菌BL21(DE3)感受态细胞,筛选表达菌株,用IPTG进行诱导表达,收集适量菌液经超声破碎后离心,取上清蛋白经SDS-PAGE电泳,蛋白质免疫印迹和活性染色分析。实验结果表明该工程菌可分泌表达一相对分子质量大约为16kD的蛋白质,与人Cu,Zn-SOD相对分子质量大小一致,表达量可达菌体可溶性蛋白的20%以上。
B.人Cu,Zn-SOD基因在酵母中的克隆与表达
将酿酒酵母交配因子(MF)-信号肽序列与hCu,Zn-SOD基因融合,首次在酿酒酵母中克隆和分泌表达了有生物活性的人Cu,Zn-SOD。酵母是真核微生物,具有不同于其他表达系统的优势:其一,与大肠杆菌不同,酵母有先进的异源蛋白折叠途径;其二,当同时使用酵母信号序列时,酵母可以分泌经正确折叠和加工的蛋白质。因此,有功能的和完全折叠的异源蛋白可以分泌到培养基中。不同于哺乳动物表达体系,酵母可以在简单培养基中快速生长。随着工业规模发酵技术的广泛应用,酵母在工业上应用于表达重要的蛋白质方面显示了极大价值。目前用酿酒酵母等真核生物细胞作为宿主表达人源SOD的研究和开发则较少。施惠娟等研究了人Cu,Zn-SOD在甲醇酵母中的表达,其表达量及酶活性均没有大肠杆菌表达量高,但由于酵母是真核表达系统,与E.coli表达系统相比,酵母表达的SOD是可溶性蛋白,且N-末端的Ala是乙酰化的,与天然的SOD结构一致,而E.coli中则没有乙酰化功能,所以酵母表达外源蛋白的生物特性更接近于天然产物,且没有内毒素污染的问题,因此用酵母表达人SOD具有很高的实用价值。人Cu,Zn-SOD蛋白为一单体,理论上相对分子质量约为16kD,大小适中,适合用酵母系统表达。
此外,由于表达蛋白的主要目的是应用于食品、化妆品生产等领域,因此在遗传背景清楚的酿酒酵母胞系统表达是很合适的;对胞内和胞外表达产物进行蛋白质免疫杂交和活性染色定位均检测到有活性的人Cu,Zn-SOD,说明有部分人源SOD分泌到胞外。
C.人Cu,Zn-SOD基因在其他表达系统中的克隆与表达
应用PCR定点突变技术把质粒pET-SOD中人Cu,Zn-SOD的Cys密码子突变为Ala密码子,再构建重组子,通过随机同源重组将突变后的人Cu,Zn-SOD整合入聚球藻并实现表达。热稳定性测定显示,重组人Cu,Zn-SOD在80℃保温30min后仍具有95%的活性,耐热能力比天然人Cu,Zn-SOD有了较大的提高。蛋白扫描结果显示目的蛋白占可溶性蛋白的3.61%。目前市场上使用的Cu,Zn-SOD基本上是动植物来源的,但不同物种的Cu,Zn-SOD具有种质特异性,特别在一级结构上差异显著,通过NCBI比较Cu,Zn-SOD的编码基因(mRNA)发现:人Cu,Zn-SOD编码基因和黑猩猩的Cu,Zn-SOD编码基因有15个碱基差异,和猕猴的Cu,Zn-SOD编码基因有22个碱基差异,故使用动植物来源的Cu,Zn-SOD存在着免疫原性等安全隐患。故近年来研究人员主要把目光投向克隆和表达的人Cu,Zn-SOD。此外自然界天然的Cu,Zn-SOD稳定性较差,所以在现实应用中受到很大的限制。研究在无毒的聚球藻中直接表达人Cu,Zn-SOD,并进一步对人Cu,Zn-SOD基因进行定点突变以提高其稳定性,为开发出一种可以直接口服、稳定而无免疫原性的新型人Cu,Zn-SOD打下基础。
7.1.3.2 微生物来源Cu,Zn-SOD基因的克隆与表达
从蜡样芽孢杆菌M22基因组DNA扩增到长1 320bp的基因片段。该片段含编码179个氨基酸残基的开放阅读框,推定蛋白序列与Bacillus anthracis Cu,Zn-SOD序列有96%同源性,其中N-端16个氨基酸残基推定为信号肽序列。将Cu,Zn-SOD编码区插入载体pET-22b(+)构建表达载体pET-22b-sod,转化E.coli BL21(DE3),IPTG诱导融合蛋白表达。SDS-PAGE显示,融合蛋白表观相对分子质量约24kD,占菌体裂解液中总蛋白的21.3%。将此表达载体转入SOD缺陷型大肠杆菌PN132,赋予了该菌株对paraquat的抗性。NBT光抑制法测定SOD活性显示,与PN132无SOD活性相比,重组子在IPTG诱导前SOD活性极低(1.79u/g),诱导后活性高达69.76u/g。非变性电泳结果显示,该酶活性不同程度地受到5mmol/L H2O2和5mmol/L KCN的抑制。
由于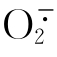 不能在生理条件下自由通过细胞膜,所以必须在其产生部位被及时清除。真核细胞中Cu,Zn-SOD主要存在于胞质内。原核生物中Cu,Zn-SOD被分泌到胞外。由于细胞外周质自身不能产生,所以胞外的Cu,Zn-SOD在抵御外界超氧自由基对机体伤害方面具有位置优势。革兰氏阳性细菌没有细胞外膜,所以Cu,Zn-SOD要么结合到细胞外膜上,要么被分泌到外部空间。从得到的B-cereusM22Cu,Zn-SOD推定氨基酸序列看,其N-端含有一个三联前导序列,即一个带正电荷的结构域、含保守的赖氨酸或精氡酸残基和一个疏水的H-结构域。再加上信号肽酶切点。这是一个典型的脂蛋白信号肽(Lipoprotein Signal Peptide)序列。该序列与芽孢杆菌属脂修饰型分泌蛋白信号肽序列有很高相似性。因此认为,B-cereusM22Cu,Zn-SOD是外泌型脂蛋白。目前还不清楚该蛋白是锚定在细胞外被还是被分泌到胞外空间。将该蛋白信号肽序列构建到绿色荧光蛋白(GFP)基因的上游,并在大肠杆菌和M22中进行表达,以便对该蛋白进行亚细胞学定位,为进一步研究其生理学功能提供依据。
不能在生理条件下自由通过细胞膜,所以必须在其产生部位被及时清除。真核细胞中Cu,Zn-SOD主要存在于胞质内。原核生物中Cu,Zn-SOD被分泌到胞外。由于细胞外周质自身不能产生,所以胞外的Cu,Zn-SOD在抵御外界超氧自由基对机体伤害方面具有位置优势。革兰氏阳性细菌没有细胞外膜,所以Cu,Zn-SOD要么结合到细胞外膜上,要么被分泌到外部空间。从得到的B-cereusM22Cu,Zn-SOD推定氨基酸序列看,其N-端含有一个三联前导序列,即一个带正电荷的结构域、含保守的赖氨酸或精氡酸残基和一个疏水的H-结构域。再加上信号肽酶切点。这是一个典型的脂蛋白信号肽(Lipoprotein Signal Peptide)序列。该序列与芽孢杆菌属脂修饰型分泌蛋白信号肽序列有很高相似性。因此认为,B-cereusM22Cu,Zn-SOD是外泌型脂蛋白。目前还不清楚该蛋白是锚定在细胞外被还是被分泌到胞外空间。将该蛋白信号肽序列构建到绿色荧光蛋白(GFP)基因的上游,并在大肠杆菌和M22中进行表达,以便对该蛋白进行亚细胞学定位,为进一步研究其生理学功能提供依据。
SOD是野桑蚕保护酶体系的重要酶类。利用RT-PCR方法克隆出野桑蚕Cu,Zn-SOD cDNA,野桑蚕Cu,Zn-SOD cDNA长497bp,开放阅读框长为465bp,从2到466位。利用蛋白分析软件EXPASY,该基因推导编码154个氨基酸,相对分子质量为15.8kD,等电点为5.78。利用PSort和TMpred进行在线分析,结果表明此蛋白质不存在信号肽和跨膜区。利用ExPASy的ScanProsite软件进行野桑蚕Cu,Zn-SOD结构和功能域分析,预测出有2个Cu,Zn-SOD的特异序列为GFHVHEFGDNT、GNAGGRIACGVI,分别位于43~53位、138~149位。前者序列中2个组氨酸分别配位结合1个铜原子;后者序列中半胱氨酸用于形成二硫键。同时预测到1个细胞黏附序列、6个N-末端酰基化位点、1个酪蛋白激酶Ⅱ(CKⅡ)磷酸化位点和1个微体C-末端导向信号。同源性及系统进化分析表明,推导出的氨基酸序列与3种果蝇的同源性平均为69.3%,与线虫为57%,各物种中与Cu/Zn结合的残基高度保守。
在野桑蚕中肠和脂肪体中分别获得了Cu,Zn-SOD基因,利用表达载体pET-28a(+)成功表达了Cu,Zn-SOD的融合蛋白,这为在野桑蚕模型中系统研究3种SOD同工酶基因结构功能、组织表达特异性、系统进化提供了有利的条件,进而为在昆虫模型上研究SOD基因的表达调控机制及其应用奠定了基础。对进一步研究鳞翅目昆虫SOD基因的表达调控机制及其应用具有重要意义。
7.1.3.3 植物来源Cu,Zn-SOD基因的克隆与表达
以水稻(Oryza Sativa)品种Cpslo17幼穗为材料,通过RT-PCR克隆了长度为698bp的编码水稻细胞质铜锌超氧化物歧化酶基因(OsCu,Zn-SOD)。序列分析表明其覆盖基因完整编码框,编码由152个氨基酸组成的蛋白,它与玉米(Zeanlize)Cu,Zn-SOD基因序列的相似率为88%。TargetP和ChloroP预测编码蛋白N-端无信号肽序列,且此编码区段有8个外显子和7个内含子。RT-PCR结果表明OsCu,Zn-SOD在水稻不同品种均有表达,但表达量存在差异,OsCu,Zn-SOD在Cpslo17的分裂旺盛的幼嫩组织如幼穗、开花期花序和未成熟种子中表达量较高,而在叶片中表达量相对较低,在愈伤组织中表达微弱。
用RT-PCR方法,从中国莲幼叶cDNA中成功扩增出Cu,Zn-SOD全长序列,并进行测序分析。该cDNA序列全长461bp,包括起始密码子和终止密码子。其中编码区段为456bp,共编码152个氨基酸,等电点为4.515。与GenBank中其他物种的Cu,Zn-SOD进行同源性比较,发现其核苷酸序列相似性高达80%~86%,氨基酸序列相似性达到89%~94%。所有已知的Cu,Zn-SOD的关键活性位点在该蛋白中均保守存在。将其与一些典型的单子叶和双子叶植物的Cu,Zn-SOD进行进化树分析,在分子生物学上为莲的系统学地位的确定提供了一个依据。
白骨壤是一种生长在热带、亚热带海岸潮间带的红树植物种类,高盐低温及海水淹浸等对其生长和分布产生重要影响。进行Cu,Zn-SOD基因的克隆研究是探讨Cu,Zn-SOD酶与白骨壤植物抗逆性关系的基础,有助于揭示白骨壤红树植物的抗逆生理机制,并为其引种驯化和扩种栽培提供依据。对重组克隆子的测序结果表明,白骨壤Cu,Zn-SOD基因片段的长度为767bp,根据与其他植物的序列进行比对得到其含4个外显子和3个内含子。4个外显子的位置依次为:1~39,171~266,582~613,701~767,总长为234bp,编码78个氨基酸,对应于水稻和玉米等植物Cu,Zn-SOD酶基因的第3,4,5,6个外显子。3个内含子的长度分别为130bp,315bp和87bp,对应于水稻和玉米等植物Cu,Zn-SOD酶基因的第3,4,5个内含子。由外显子核苷酸序列推导的氨基酸序列与水稻(GenBank编号:L19435)、玉米(GenBank编号:M15175)、红薯(GenBank编号:L36229)和杨树(GenBank编号:AF016893)的Cu,Zn-SOD基因编码的相应氨基酸序列相似,同源性分别为83.3%,84.6%,84.6%和87.2%,说明Cu,Zn-SOD基因高度保守。
对不同植物间克隆的Cu,Zn-SOD基因序列进行比较,可以发现不同植物间除其外显子序列具有较强的保守性外,其内含子的位置和数目也具有很强的保守性,说明不同植物间Cu,Zn-SOD基因结构的稳定性。但是不同植物间Cu,Zn-SOD基因的内含子序列差别很大,长度各异。内含子序列中含有对基因表达起调控作用的元件,这种序列和长度上的不同可能导致不同植物对环境信号和逆境的差异性反应。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。










