



I
我们现在已经讨论完毕就目前所知的并已充分确定的关于稳态的几个方面和我们对躯体天然防卫的若干问题以及结构和机能的安全系数。要想进一步深入观察机体所造就的、经得起环境剧烈冲击的弹性与耐力,就得考察稳定性赖以实现的那些途径。
作为试图了解液床中稳定状态的管理方式的一个基础,我们必须记住神经系统机能的概要。神经系统按照其机能可分为两大部分:一个是个体与外环境相联系的对外反应的部分;另一个是作用于内脏即主要是管理内环境的对内部分。我们的兴趣特别在于后一个部分,但在正常生活中这两个部分是分不开的。
II
我们都熟悉脑和脊髓的外观以及它们在颅骨和脊椎骨内各自的位置。我们也知道神经纤维把脑和脊髓与体表上每个点以及我们能运动的每个肌肉连结起来。从体表传入的纤维即所谓“感觉纤维”所接受的刺激是通过感觉器官,也就是“感受器”或“外部感受器”(exteroceptor)(以区别于内部感觉器官)来传递的。这些外部感受器通常是细胞或细胞群或细胞层,它们能分别感受不同类型的外界刺激。这些感受器计包括接受触觉刺激的触觉小体,接受冷热的温觉末梢,接受空气中化学物质刺激的鼻内嗅区,位于舌上的接受溶于水中的某些化学物质刺激的味蕾,感受通过骨性杠杆所传导的空气震动的内耳,还有对光波具有敏锐感觉的眼。某些感受器要求与刺激因素直接接触,例如触觉和味觉感受器,其他如嗅、听及视觉部分则可感受躯体以外不同程度距离的变化,因而被称为“远距离感受器”(distance receptor)。通过这些体表的敏感器官,我们得以熟知周围物体的一切种类和状态,从一块布的质地直到亿万光年之遥的星球的特征。
传入神经纤维把感觉的神经冲动从每个外部感受器传导到脑或脊髓——即“中枢神经系统”。在外界因素刺激感受器的时刻,这些神经纤维报告刺激时间以及刺激的程度和特点。
从中枢神经系统发出一些神经纤维到操纵骨骼活动的肌肉,也分布到另外一些不附着于骨骼的面部肌肉。接受这些“运动神经”支配的肌肉称为“效应器”或“效应器官”。当然,“运动神经”除非能够使肌肉活动,否则是无用的,而肌肉如果不受神经冲动的支配而运动也会同样变成无用的,就像瘫痪病人那个样子。因此,自然应该把神经系统的对外作用部分看做是一种神经——肌肉组织(neuro-muscular organization)。
脑和脊髓是由极为错综复杂的神经束装置组成的,这些神经束能把来自外部感受器的输入通路上的每一个站与通向效应器的输出通路上的每一个站之间连结起来。因而个体对所接受的一种刺激(例如左肩部的痒感),或反映复杂的外界状态的一组刺激(例如一个脱缰的马)都能够发生特定的调节反应,从而使所产生的行动适应其需要。
III
最简单的反应就是反射,即机体对感受器传入刺激发生反应,立刻发出运动冲动而并引起肌肉收缩。喷嚏、咳嗽、眨眼以及保持姿势,都是反射的例证。这些反射通常都具有保护作用。这些动作可以说都是不随意的,而且不伴有任何意识的活动。还有更复杂一些的反应是与感觉和情绪相关联的,诸如笑、哭,还有种种典型表现的愤怒与恐惧。这些反应和反射一样都不是学会的;因为它们能在人生的早期即有所表现,作为模仿动作来说是为时过早了。负责管理反射活动的部分主要在脊髓和脑的下部(即延髓和中脑,见图34),该部紧连着脊髓。这些反应属于本能类型,近似于反射,其中枢位于脑的底部。例如,愤怒的生理表现,如巴尔德所指出的,在间脑以上所有部分都被切除后仍能得到相当充分的显示。这种现象在进一步切除间脑后立即消失。
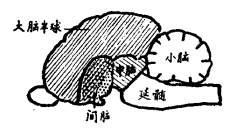
图34:哺乳类脑的正中切面。CH,大脑半球;D,间脑(用点标出部分);M,中脑;Md,延髓;Cb,小脑。
位于大脑半球外层的大脑皮质,是负责联想性记忆和一切对周围环境的肌肉反应进行复杂调节的一种器官(见图34)。在中枢神经系统的其他部分我们和低等动物可能没有太大的不同,但在大脑半球中,我们有几乎无与伦比的差异。与任何低等动物相比,人的感受器和肌肉效应器的神经联系在这个区域中是最丰富和最细致的,而且在感受器与效应器的中枢之间相互联系也复杂得多。人与其他哺乳动物相比之所以具有更高的智慧和改造外部世界的非凡能力,就是建立在这种大脑皮质神经联系的精巧构造基础之上的。
IV
有机的生物体必然要遇到的基本问题,即继续生存和种族繁殖,要求两个基本的活动:运动与捉拿(grasping)。这类活动一向是以各种不同的巧妙的方式进行着的。照例,鱼类以上的脊椎动物,运动是四肢的一种机能,即使鱼类也是利用与四肢相当的器官即鳍和尾部来达到运动的目的。在鸟类,在其进化的实验过程中,曾经力图改造其前肢以实现其空中运动的目的。但把两对肢体都用于运动,就使多数脊椎动物把捉拿的机能留给了牙齿,鸟类则用喙,象则用鼻。这自然反映了器官的专门用途或双重用途;蛙用它的短腿抓取,熊站着抱物,松鼠以其前爪抓核桃,而猿猴类则有一双类似手的脚,不仅用来行走也能用以捕捉。然而在人类,充分完善的机能分化达到了这样的地步,即后肢专司运动而前肢和手专司捉拿。手能执行一切方式的动作,这种突出机能的发展显然是与前脑和大脑皮质的复杂结构的发展分不开的。手创造了工具和器械——镐,锯,刷子,手术刀,车床,汽锤以及不论其他什么东西!这些东西惊人地提高了手工操作的强度和精巧度。工具和器械又反过来用于创造能够大大扩展我们感受器的作用范围的装置——例如用于观察细微物体的显微镜和用于观察远方物体的望远镜以及能从远距离收听大气中精微的电脉冲的无线电收音机和扩大器。
脑脊髓神经系统精巧地配备着敏感的外部感受器和一批能达到所要求的任何角度以及几乎向任何方向操纵许多骨骼的肌肉群,这个神经系统的作用可使机体通过劳动、奔跑和战斗来改变外界环境或改变本身在此环境中的位置。这些对外的活动可以专门叫做“外效应”(exterofective)活动,而所谓“随意神经系统”可以很恰当地称为神经系统的外效应部分(exterofective division)。但我们已经知道,外效应活动必然会使内环境也产生相应的变化,如血糖的利用,酸性废物和过多的热向血流内排放等。在这种状态下,“不随意神经系统”通过对心脏和其他内脏的肌肉、腺体的影响而发挥作用以维护内环境使之适合随意运动的继续。由于不随意神经系统这种对内机能的存在,有理由称它为内效应(interofective)系统[1]。现在我们就来讨论这个系统的组成情况。
V
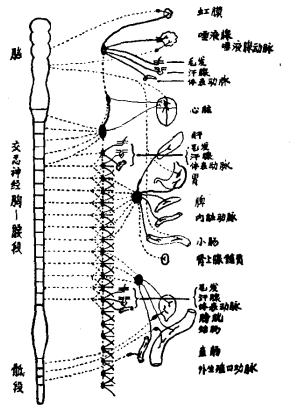
图35:自主神经系统分布略图。左侧为脑和脊髓部分。图中未画出躯体系统的神经。节前纤维用虚线表示,节后纤维用实线表示。详见正文。(摘自Bard〔after Cannon〕,“Foundations of Experimental Psychology.”Courtesy of the Clark University Press.)
神经系统的内效应部分除称为“不随意”系统外,又称为“植物性”或“自主”神经系统,称为“植物性”是因为,它与运动、捕捉这种动物性机能相比,在更大程度上与机体的营养有关;而称为“自主”是由于它能自动地动作而不靠大脑皮质的命令。自主神经系统的任务是调节内脏机能以适应机体整体的需要。我们必须知道它所具有的一些特性以便能够理解后面将要提到的一些实验的意义。
首先,由自主神经纤维所支配的内脏部分是平滑肌(“平滑”一词是用来区别于附在骨骼上的横纹肌的)和腺体。平滑肌细胞可见于毛发根部、血管的被膜、环绕肺中细支气管的周围部分以及像胃、肠、膀胱、子宫这一类空腔脏器的壁层。腺体包括唾液腺、胃腺、胰腺和肝这样一类腺体。
自主神经系统的神经纤维大部分是输出的,它们从中枢神经系统的三个区段向外伸出:一是从脑部这个区段向外伸出;再是从脊髓向上、下肢发出神经的那个区段向外伸出;三是从脊髓下段发出支配下肢神经的脊髓段向外伸出。自主神经系统的这三个区段分别称为脑段,胸—腰(或交感)段,和骶段(见图35)。
在脑和脊髓发出的神经纤维与它们所支配的内脏之间,恒有一种位于其间的周围神经细胞(神经元),这些细胞及其纤维是直接作用于内脏的装置。这些神经细胞体通常集合成小的结节称为神经节(ganglion)。因此把近端神经纤维称为“节前纤维”,而其远心段则称为“节后纤维”(见图35)。
内脏的平滑肌和腺体所具有的特点是:支配它们的神经纤维有两个来源,一方面来自头端或尾端的一段(即脑段或骶段),另一方面来自中间段(交感神经)。正如我们所知道的,心脏既由属于脑段的迷走神经支配,同时也由来自交感部分(即胸腰段)的加速纤维支配。
支配任何一个内脏的来自两个节段的神经纤维,其所产生的效应照例是互相拮抗的。迷走神经作用是抑制心率,交感神经则使它加快;迷走神经增加胃部肌肉中等程度的收缩也就是增加其张力,交感神经则使之减少。还可举出许多其他例证。
脑段和骶段的特点是,它们的节前纤维在得到周围神经元的接替之前就进入或接近所支配的内脏。换言之,这些周围神经元的神经节和节后纤维位于或接近其效应器官(见图35)。这和交感神经中的情况完全不同。
VI
除了腹主动脉的一些大的分支附近有支配腹腔内脏的专有的神经节外,交感神经系统周围的神经细胞所组成的神经节分别位于脊柱两旁的两条神经链上,上自位于颈部的颈上神经节,下到骨盆中的合体(coalesced)神经节。在胸、腹腔内这些神经节位于胸、腹腔的后方,紧靠椎体。它们由来自脊髓的、依次在各脊椎间发出的节前纤维连结在一起,纤维进入最近的神经节,与其中的周围神经元相连结,然后分别向下或向上到达神经链上的其他神经节,与其中各自的神经元连结起来。这种安排提供了一个具有广泛接触的高效能的装置,从而增加了神经分布的通路。兰生(Ranson)和毕林斯利(Billingsley)曾在猫身上计算了一个颈交感神经干(由节前纤维组成)中节前纤维的数目及其颈上神经节中细胞体的数目,他们报告这二者之比为1∶32。显然,如果普遍存在着类似的关系,那么由于任何单独的节前纤维与较多的终末神经元相连结,因而它能传递具有广泛效应的神经冲动。而在胸、腹部,几个神经节之间由节前纤维搭桥形成的连结,将使这些搭桥纤维能支配所有这些神经节所支配的区域。
刚才提到的搭桥纤维也就是连结纤维,能在上下一定距离内伸到脊髓每一侧的神经节链上,因而彼此重叠。其结果就是通过交感神经通路释放出一个广泛扩散的神经冲动,与脑段与骶段机能相对比,可以看出后者对一些特定器官所发出的冲动有明确针对性和局限性。
交感神经的结构具有扩散性效应这种看法得到一般观察的支持。例如当一只猫接受兴奋或寒冷的刺激——即引起交感神经兴奋状态时,从头部到尾巴都发生了竖毛现象。更深入的观察发现,这种全身竖毛的现象标志着种种大的系统有其他的一般效应,例如,整个消化道活动的抑制以及全身各部分血管的收缩。另一方面,脑段(副交感中枢)并非必然地同时去执行一些不同的职能,而是可以分别地来完成的。因此,当眼睛接受十分强烈的光刺激时所产生的冲动是沿脑神经的植物神经的通路传递的并引起了虹膜的收缩,这个冲动并不必然地伴随另外一些会引起唾液分泌增加或心率减慢或使胃肠道张力增加的神经冲动。脑段的这些效应,正如分布在一些器官的节前纤维的效应一样是各自不同的。
交感神经冲动能引起肾上腺髓质分泌出肾上腺素这一事实,同样肯定了交感神经的结构是供神经冲动作扩散性释放这样一种观点的。这种物质(肾上腺素)如果注射到血流中,它照例会在交感神经所支配的器官中产生出等同于神经冲动所引起的效应。由于分泌出来的肾上腺素在血流中的广泛分布,交感神经系统即使没有本身神经纤维的组成方式所带来的扩散性效应,也能由于肾上腺素的作用而具有这样的效应。但是这两个因素——神经冲动和血循环中模拟神经效应的化学物质,一般作为交感—肾上腺系统而共同起作用,从而在整个机体的平滑肌和腺体中产生广泛的变化。
这里产生了这样的问题:在特殊情况下,过多的肾上腺素分泌到血循环中,是否该血中含量足以在事实上影响到内脏呢?有许多观察能对这个问题给出肯定的答复。在前面的讨论中,我们曾经指出了在低血糖及寒冷刺激下,肾上腺以自然方式分泌的肾上腺素对去神经心脏的效应(见图18和28)。类似的证明曾见于去神经的唾液腺、虹膜和肾脏,也见于通过交叉循环——即将一个动物的肾上腺静脉连接到另一个动物的大循环上——所得到的结果。通过所有这些实验方法得以证明,分泌到血液中的肾上腺素浓度对用于试验目的的远隔器官足以产生有效的作用。
虽然说肾上腺素通过血液传送而能与交感神经冲动协同产生与交感神经冲动所引起的同样的效应,但又如何证明肾上腺素的协同作用的有效性呢?首先,有若干指征说明血循环中肾上腺素能够延长交感神经作用的效应。布利顿和我曾报告过去神经心脏心率的持续加速现象,即在一分钟的兴奋后加速现象持续了近半小时,虽然该动物在短暂的不安后仍然安静地卧在软垫上(见图36)。
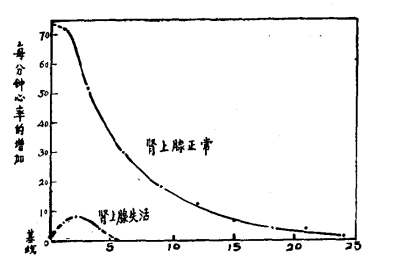 图36:在肾上腺正常的情况下,动物(猫)用狗吠刺激1分钟使之兴奋后心率长时间增加的持续现象。在肾上腺失活后,虽然对猫的刺激加倍,心率只有轻度增加并较快复原。在两种情况下,猫经刺激后都迅速从笼中移出,恢复期间一直安静地卧在软垫上。
图36:在肾上腺正常的情况下,动物(猫)用狗吠刺激1分钟使之兴奋后心率长时间增加的持续现象。在肾上腺失活后,虽然对猫的刺激加倍,心率只有轻度增加并较快复原。在两种情况下,猫经刺激后都迅速从笼中移出,恢复期间一直安静地卧在软垫上。
可见,当情况需要作用延长时,分泌的肾上腺素就可以起作用。其次,一些观察结果表明,分泌的肾上腺素所产生的效应在某些方面还远超过交感神经冲动单独所能引起的作用。例如窒息、反射刺激、正常的情绪兴奋以及切除大脑半球皮质后所出现的假怒现象,都伴有高血糖现象。这种变化在切断肝脏神经而保留了肾上腺的动物中与正常动物所出现的程度完全一致。相反,如果肾上腺被切除或失去活性而保留了肝神经,则在同样实验条件下,可看到高血糖现象减少或完全不发生。十分可能的是,肾上腺素加快血凝过程和代谢速度的作用,与其消除肌肉疲劳后果的独特作用相同,不依靠交感神经冲动的任何协同作用。
VII
调节活动能否实现,是与内脏的双重神经支配以及与任何脏器中这两种神经在作用上照例是相互拮抗的这种重要事实密切相关。通过交感部分的广泛扩散效应和脑—骶部分一些神经特有的拮抗效应保证了局部的和暂时的变化的多样性。通过交感神经的适度的或强度的(增加或降低)改变,可以使所有内脏向一方或另一方同时受到影响。而通过直接到达某内脏的与交感部分相对立的脑段或骶段特定神经强度的增减,可使特定的内脏向着一个方向或另一方向有所不同地受到影响。举例说,在情绪激动时,由于交感神经所产生的对内脏的复合效应之中的部分效应,心率可以加快,但是,心率加快也可由于对迷走神经抑制的减弱而不是广泛地涉及其他内脏,如同在肌肉运动开始阶段中所见到的那样。交感神经犹如高音或低音踏板,普遍调节所有的音符;而脑部和骶部的神经支配就像各个琴键。如果我们考虑到紧急情况下交感系统机能以多种多样的方式从整体上来保护机体的整体,交感神经具有同时作用和统一作用这种结构安排的重要性就更加明显了。
VIII
用“不随意”和“自主”这样的名词来表现神经系统的内效应部分的特点,其含意是将它与操纵骨骼肌的系统区分开来。我们不能靠意志的作用来抑制胃肠运动、减慢心脏搏动或从肝内释放糖。但是,所有这些以及更多的变化都会在适当的时机通过自主神经冲动的作用来完成。自主神经系统中的反应明显地与简单反射有着密切关联,这些简单反射是迅速的、先天的、不随意的和具有目的性的。因此,通过考察自主神经系统每一个机能发生作用的特定环境,以及每一个机能所产生的特定效应,我们可能对自主神经系统的机能得到深入的了解。下面的提要和我在1914年所提出的设想是一致的。
骶段的机能主要是对那些周期性充盈的空腔器官加以排空,因而骶部自主神经冲动引起直肠和远段结肠的收缩,并引起膀胱、可能还有其他容纳器官的收缩。最为人们所熟知的例子是,具有张力收缩的内脏,由于其中积存的内容物而发生扩张,从而反射性地引起了收缩效应。虽然这种对膨胀引起反应的反射可能是骶段作用的最常见的方式,但能引起反射的还有其他方式的刺激。例如巨大的情绪冲动——恐惧,它所引起的神经冲动可通过交感部分释放,也可能通过骶神经通路释放。膀胱和低位大肠在强烈兴奋中发生不自主的排泄,这在人和其他动物中是常见的现象。而特殊的动感情状态则能显著影响生殖器官的机能。由于在某些情况下骨盆区的空腔器官必须通过横纹肌构成的括约肌来向外排放,这些器官的排空在某种程度上有大脑皮质的即随意的控制。通过腹肌的随意收缩也能促进排空过程,但从其本质特征来说,骶段机能仍是自主的。事实上,如在世界大战中伤员身上所看到的,这些机能中枢所在的脊髓段与脑部间的联系完全被切断之后,仍然能够执行其机能。
脑段的机能,和骶段一样,概括起来可以这样说:它们主要是一组反射,具有防护、保存和加强的作用。脑段通过缩小瞳孔来保护视网膜以防过强的光线。它通过唾液和胃液的分泌并通过给消化管壁肌肉带来周期性收缩所需的张力为全身活动所需的能源物质的特定消化与吸收提供了根本依据。它通过迷走神经对胰岛素分泌的控制,可以对肝糖原的储存发生作用。通过迷走神经的紧张使心率减慢来保证心肌的休息及其恢复的时间,从这里又找到了脑自主神经作用保护性意义的进一步证明。
脑段同骶段一样,不仅有缩小瞳孔这一类非复杂反射,而且还有联系到情感状态的活动。食物的美味和香味引起所谓消化液的“精神性”分泌,还引起胃肠的张力性收缩。此外,这两个部分——骶段与脑段——的相似点是:它们都在很大程度上受横纹肌运动的影响。正如膀胱和直肠的收缩能受大脑皮质神经冲动的加强或抑制一样,瞳孔的对光或对距离的反应也能被随意动作而引起改变。实际上,作为一般规律,骶段和脑段的活动比起交感神经来更多地包括了大脑脊髓神经系统的协同作用,因为它们与横纹肌所围绕的外部管道口有更多的关系。
IX
作为对内脏活动起到影响的内效应系统,自主神经必然要密切地涉及保持机体内部系统的稳定性与恒定性,我们把机体内部这种稳定状态称之为稳态。事实上,自主神经系统的三个部分的许多机能可以合理地被看成就是为了维持机体的稳态。骶段主掌周期性充盈的空腔器官的一组排空反射,并通过排除废物和排泄那些妨碍或限制个体行动自由的蓄积物来保持恒定性。另外也参与保证了种族的延续。脑段则执行一组具有防卫和保护作用的反射,更明显地涉及机体整体性的保持。就其主要作用范围——调节食物的消化与吸收——而言,对维持稳定状态有着根本的重要性。
然而,内效应系统的脑段和骶段只是以间接的和多少有点远隔的方式来确保恒定状态。能迅速和直接地防止内环境剧烈变化的还是中段即自主神经系的胸腰段。胸腰段的作用对稳态是如此重要,值得进行专门的、详尽的考察。
参考文献
Bard. Am. Journ, Physiol., 1928, lxxxiv, 490.
Cannon. Am. Journ. Psychol., 1914, xxv, 256.
Cannon. The Autonomic Nervous System, an Interpretation, The Lancet, 1930, i, 1109.
Cannon and Britton. Am. Journ. Physiol., 1927, lxxix, 433.
Ranson and Billingsley. Journ. Comp. Neurol., 1918, xxix, 305.
[1] “外效应”、“内效应”皆为著者所命名。内效应系统又译为内环境调节系统。——译者
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。










