



第一节 白细胞分化抗原
一、概述
白细胞分化抗原(leukocyte differentiation antigen,LDA)是白细胞(还包括血小板、血管内皮细胞等)在分化成熟为不同谱系(lin-eage)和分化不同阶段以及活化过程中,出现或消失的细胞表面标记。它们大都是穿膜的蛋白或糖蛋白,含胞膜外区、穿膜区和胞浆区;有些白细胞分化抗原是以糖基磷脂酰肌醇(glyco-sylphosphatidylinositol,GPI)连接方式“锚”在细胞膜上,少数白细胞分化抗原是碳水化合物半抗原。
白细胞分化抗原种类繁多,分布广泛,除表达于白细胞之外,还广泛分布于不同分化阶段的红细胞系、巨核细胞/血小板谱系和非造血细胞(如血管内皮细胞、成纤维细胞、上皮细胞、神经内分泌细胞等)表面。
白细胞分化抗原参与机体重要的生理和病理过程:(1)免疫应答过程中免疫细胞的相互识别,免疫细胞抗原识别、活化、增殖和分化,免疫效应功能的发挥;(2)造血细胞的分化和造血过程的调控;(3)炎症发生;(4)细胞的迁移如肿瘤细胞的转移等。本章仅介绍参与免疫细胞识别、信号转导以及活化与效应的CD分子。
早期各实验室多借助自制的特异性抗体对白细胞分化抗原进行分析和鉴定,故同一分化抗原可能有不同命名。20世纪80年代初以来,由于单克隆抗体,分子克隆、基因转染细胞系等技术在白细胞分化抗原研究中得到广泛深入的应用,有关白细胞分化抗原的研究和应用进展相当迅速。在世界卫生组织(W HO)和国际免疫学会联合会(IU IS)的组织下,自1982年至1993年已先后举行了五次有关人类白细胞分化抗原的国际协作组会议(Interna-tionalworkshop on human leukocyte differentiation antigens),并应用以单克隆抗体鉴定为主的聚类分析法,将识别同一分化抗原的来自不同实验室的单克隆抗体归为一个分化群,简称CD(cluster of differentiation),以CD代替以往的命名。迄今,人CD的序号已从CD1命名至CD339。在许多场合下,抗体及其识别的相应抗原都用同一个CD序号。
二、参与T细胞抗原识别与活化的CD分子
T细胞是一类重要的免疫活性细胞,除直接介导细胞免疫功能外,对机体免疫应答的调节起关键作用。T淋巴细胞本身的识别活化及效应功能的发挥,不仅与外来抗原、丝裂原和多种细胞因子密切相关,而且有赖于T细胞相互之间、T细胞与抗原递呈细胞(APC)之间以及T细胞与靶细胞之间的直接接触。T淋巴细胞识别抗原的受体是T细胞受体(T cell receptor,TCR)与CD3所组成的复合物(TCR-CD3)。在识别过程中还有赖于抗原非特异性的其他细胞表面分子的辅助,这些辅助分子(accessory molecules)主要包括CD4、CD8、CD2、CD28、CD40L、CD58、CD80、CD86和CD152等(图8-1)。
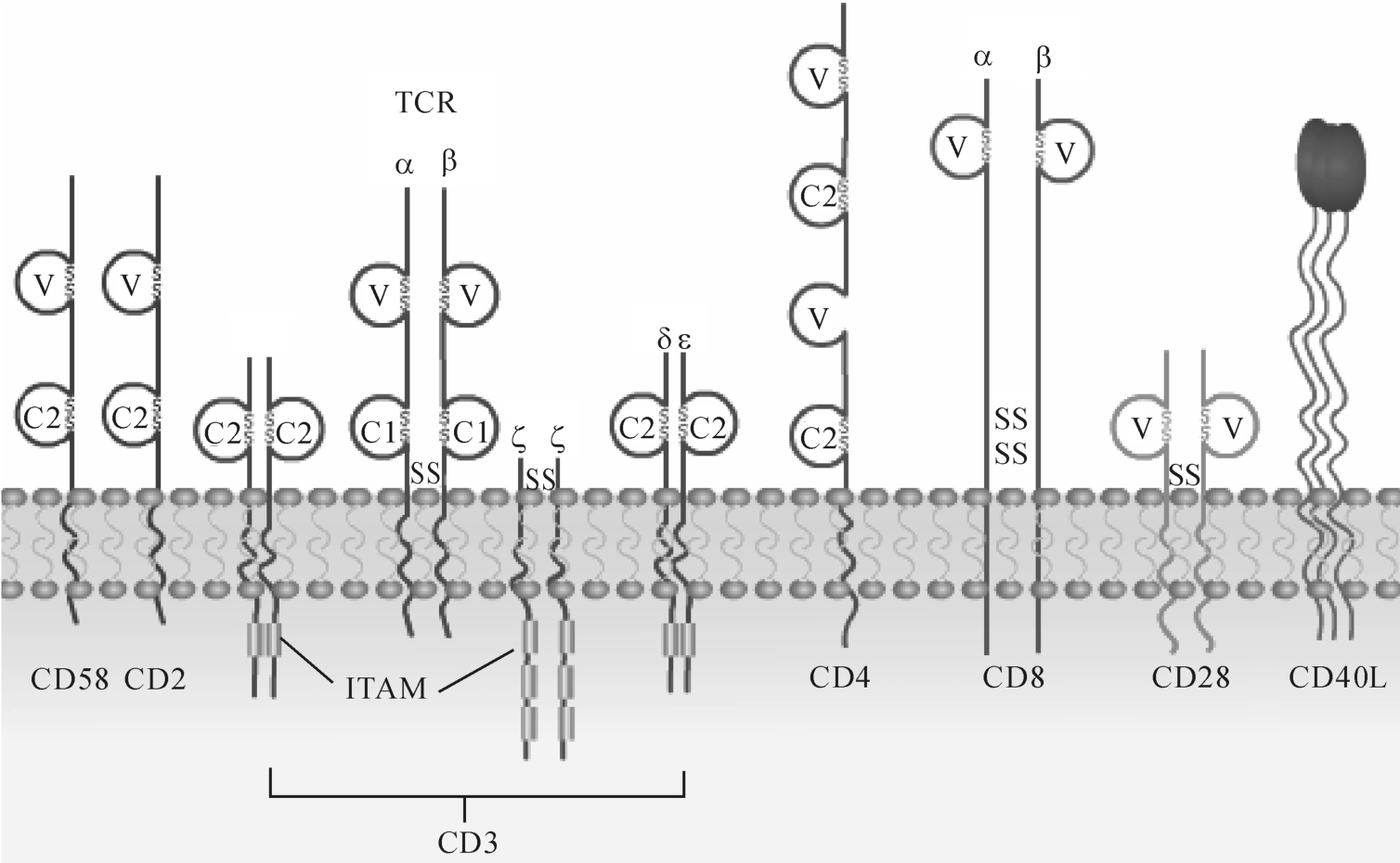
图8-1 参与T细胞抗原识别与活化的CD分子
1.CD3
CD3由γ、δ、ε、ζ、η五种肽链组成,通过盐桥与T细胞受体TCR形成TCR-CD3复合体,分布于所有成熟T细胞和部分胸腺细胞表面。
CD3的主要功能是转导TCR特异性识别抗原所产生的活化信号,促进T细胞活化。CD3分子胞浆区含免疫受体酪氨酸活化基序(immunoreceptor tyrosine-based activation motif,ITAM),TCR识别或结合由M HC分子递呈的抗原肽后,导致ITAM所含酪氨酸磷酸化,通过活化相关激酶,将识别信号转入T细胞内(图8-2)。CD3是参与TCR信号转导的关键分子,CD3肽链缺陷或缺失,可导致T细胞活化缺陷。
2.CD4
CD4为单链跨膜糖蛋白,属免疫球蛋白超家族(IgSF)成员,分布于胸腺细胞和成熟TH细胞,也存在于巨噬细胞、脑细胞。在外周血和淋巴器官中,CD4+T细胞主要为辅助性T细胞(helper T cell,Th)。功能:(1)作为TH与APC之间的黏附分子,CD4/M HC-II类。(2)信号转导作用:细胞内传导。CD4分子也是人类免疫缺陷病毒(H IV)受体。
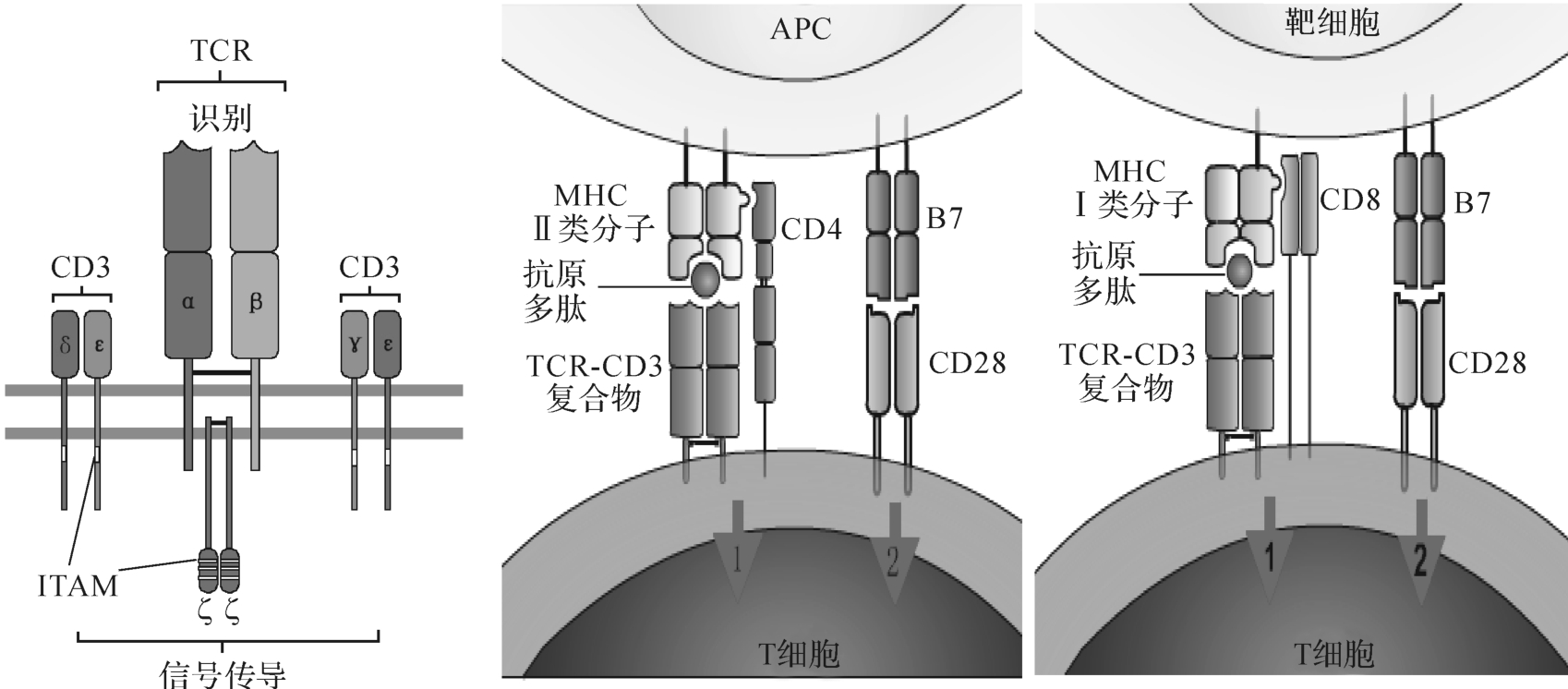
图8-2 活化信号
3.CD8
CD8也属IgSF成员,分布于部分T细胞、胸腺细胞和NK细胞表面,通常作为判别T细胞的表面标志。功能:(1)介导细胞间黏附作用:CD8与M HC-I类结合,激活CTL。(2)信号传导:CD8-M HC-I结合,启动T细胞免疫应答。
4.CD28与CD80(B7-1)/CD86(B7-2)
CD28分子乃借二硫键相连的同源二聚体,属IgSF成员。在外周血淋巴细胞中,几乎所有CD4+T细胞和50% CD8+T细胞表达CD28。此外,浆细胞和部分活化B细胞也可表达CD28。一般而言,活化T细胞CD28表达水平升高。
CD28分子胞浆区可与多种信号分子相连,能转导T细胞活化的共刺激信号。CD28的配体是表达于B细胞和APC表面的B7家族分子,包括CD80(B7-1)和CD86(B7-2)。CD28/B7-1、B7-2是一组最重要的共刺激分子,它们之间结合提供T细胞活化所必需的共刺激信号,即第二信号。
5.CD2/CD58
图8-3 E花环
CD2又称淋巴细胞功能相关抗原2(lymphocyte function associated antigen 2,LFA-2)或绵羊红细胞受体(sheep red cell receptor,SRBC),表达于T细胞、胸腺细胞和NK细胞等。人CD2的配基是CD58(LFA-3)分子,二者结构相似,且均属IgSF成员。
CD58分布较广,包括多种血细胞和某些非造血细胞。CD2与CD58结合能增强T细胞与APC或靶细胞间黏附,促进T细胞对抗原识别和CD2所介导的信号转导。此外,人T细胞还能通过CD2与SRBC表面的CD58类似物结合形成花环,称为E花环(图8-3),可用于体外检测和分离T细胞。
三、参与B细胞识别Ag与活化的CD分子
B细胞抗原受体(BCR、Sm Ig)、CD19/CD21/CD81、CD40与CD40L等(图8-4)。
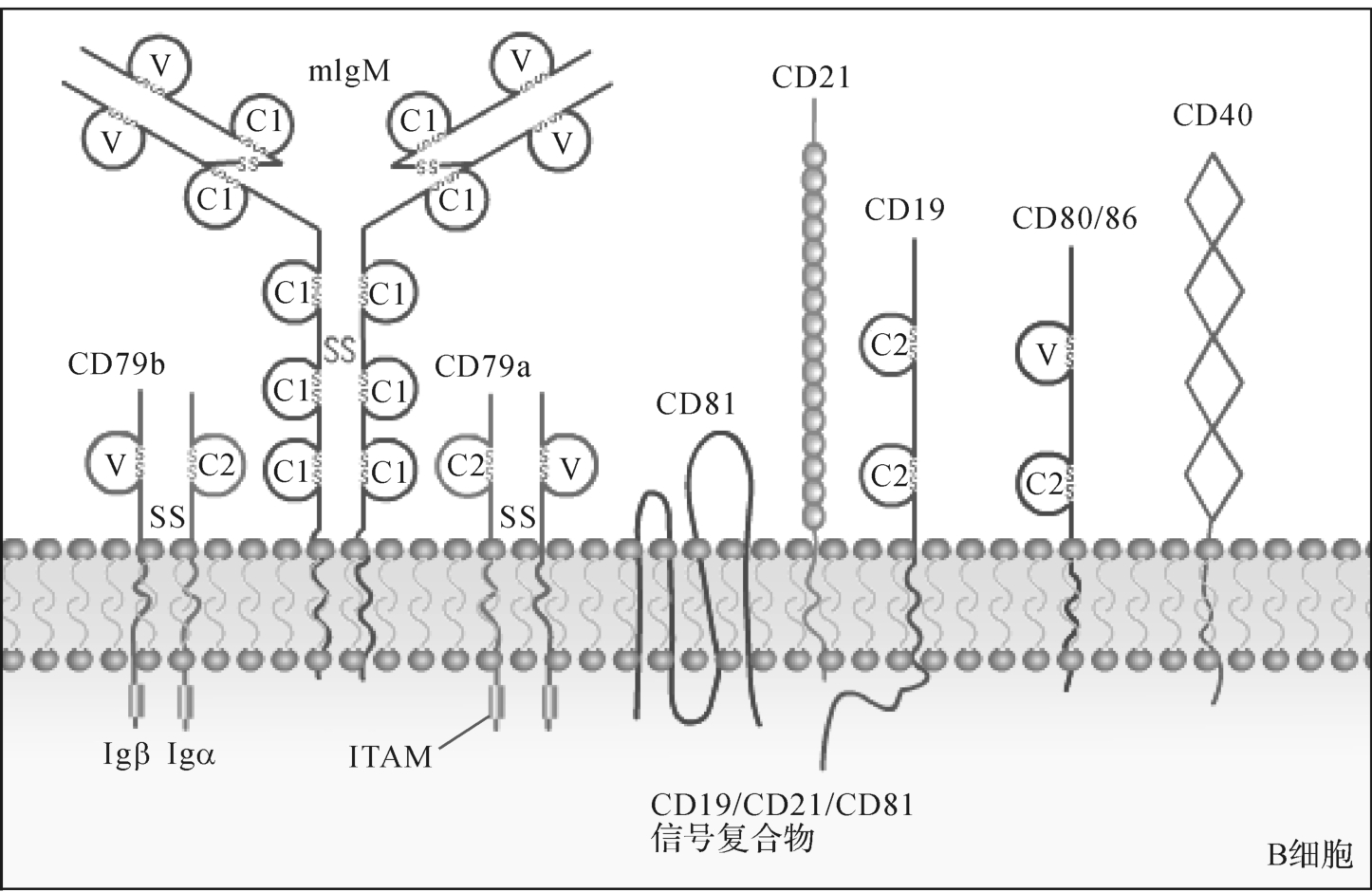
图8-4 参与B细胞抗原识别与活化的CD分子
1.B细胞抗原受体(BCR、Sm Ig)
B细胞抗原受体是B细胞特异性应答的关键分子(图8-5)。BCR特异性识别并结合抗原。BCR也有两种辅助成分,即Ig-I(CD79a)和Ig-I(CD79b)。在人类B细胞中,与m IgM相关的Igα和Igβ分别为47kDa和37kDa糖蛋白,属于免疫球蛋白超家族成员,通过非共价键成为BCR-Igα/Igβ复合体。Igα和Igβ胞膜外区氨基端处均有一个Ig样结构域。Igα和Igβ均可作为蛋白酪氨酸激酶的底物,可能与BCR信号转导有关,因为m IgM和m IgD胞浆区只有3个氨基酸(KVK),不可能单独把胞膜外的刺激信号传递到细胞内。
2.CD19/CD21/CD81
CD19、CD21、CD81构成的复合物是B细胞活化的共受体,通过CD19分子胞浆区与多种激酶的结合,能加强跨膜信号转导,促进B细胞活化。CD19/CD21/CD81信号复合物可调节BCR活化的阈值,其中CD21(CR2)借助补体C3片段而介导CD19与BCR交联,从而促进B细胞活化,这对B细胞初次应答尤为重要(图8-6)。
CD19分布于除浆细胞外不同发育阶段的B细胞表面,是鉴定B细胞的重要标志之一。CD21是CR2、C3dR、EB病毒的受体,仅表达于静止的成熟的B细胞表面,B细胞一旦活化即消失,是B细胞的重要标志。CD81广泛分布于B细胞、T细胞、巨噬细胞、树突状细胞、NK细胞和嗜酸粒细胞表面。CD81是丙型肝炎病毒(HCV)受体,可能参与HBV感染。
3.CD40与CD40L
CD40分子属肿瘤坏死因子超家族,主要分布于B细胞、树突状细胞以及某些上皮细胞、内皮细胞、成纤维细胞和活化的单核细胞表面。

图8-5 B细胞抗原受体结构示意图
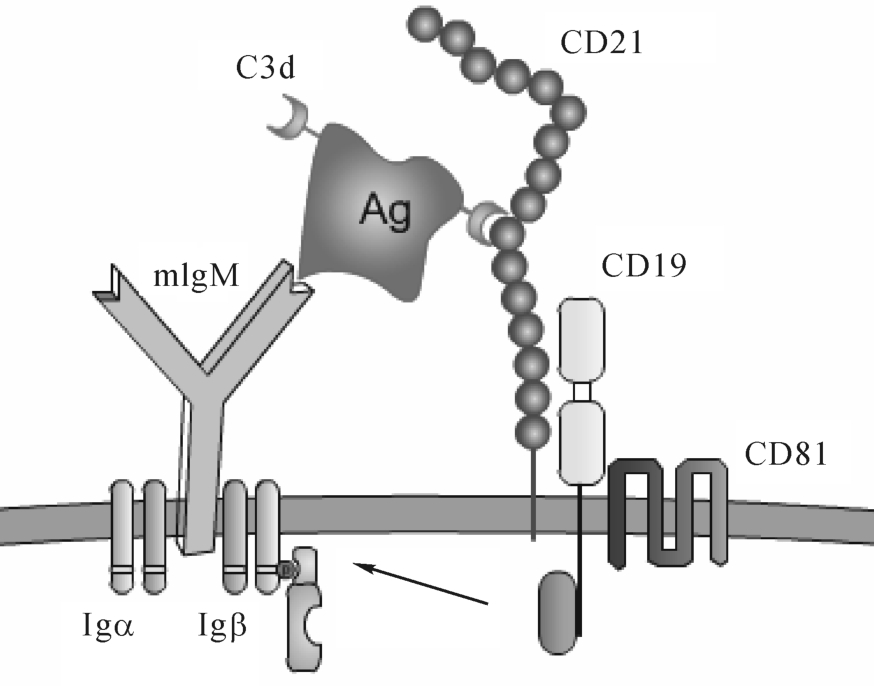
图8-6 B细胞信号复合物示意图
CD40L即CD40配体,属IgSF家族成员。人CD40L主要表达在活化CD4+T细胞、部分CD8+T细胞和γδT细胞表面。CD40L与B细胞表面CD40结合是B细胞再次免疫应答和生发中心形成的必要条件。T细胞表面CD40L与B细胞表面CD40结合,能提供B细胞活化所需的共刺激信号,这是B细胞对TD抗原产生应答的重要条件。CD40L也能激活单核/巨噬细胞。CD40与CD40L的相互作用还参与淋巴细胞发育的阴性选择过程和外周免疫耐受的形成。此外,CD40L还表达于活化的嗜碱粒细胞、肥大细胞、NK细胞、单核细胞以及活化B细胞表面。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。










